
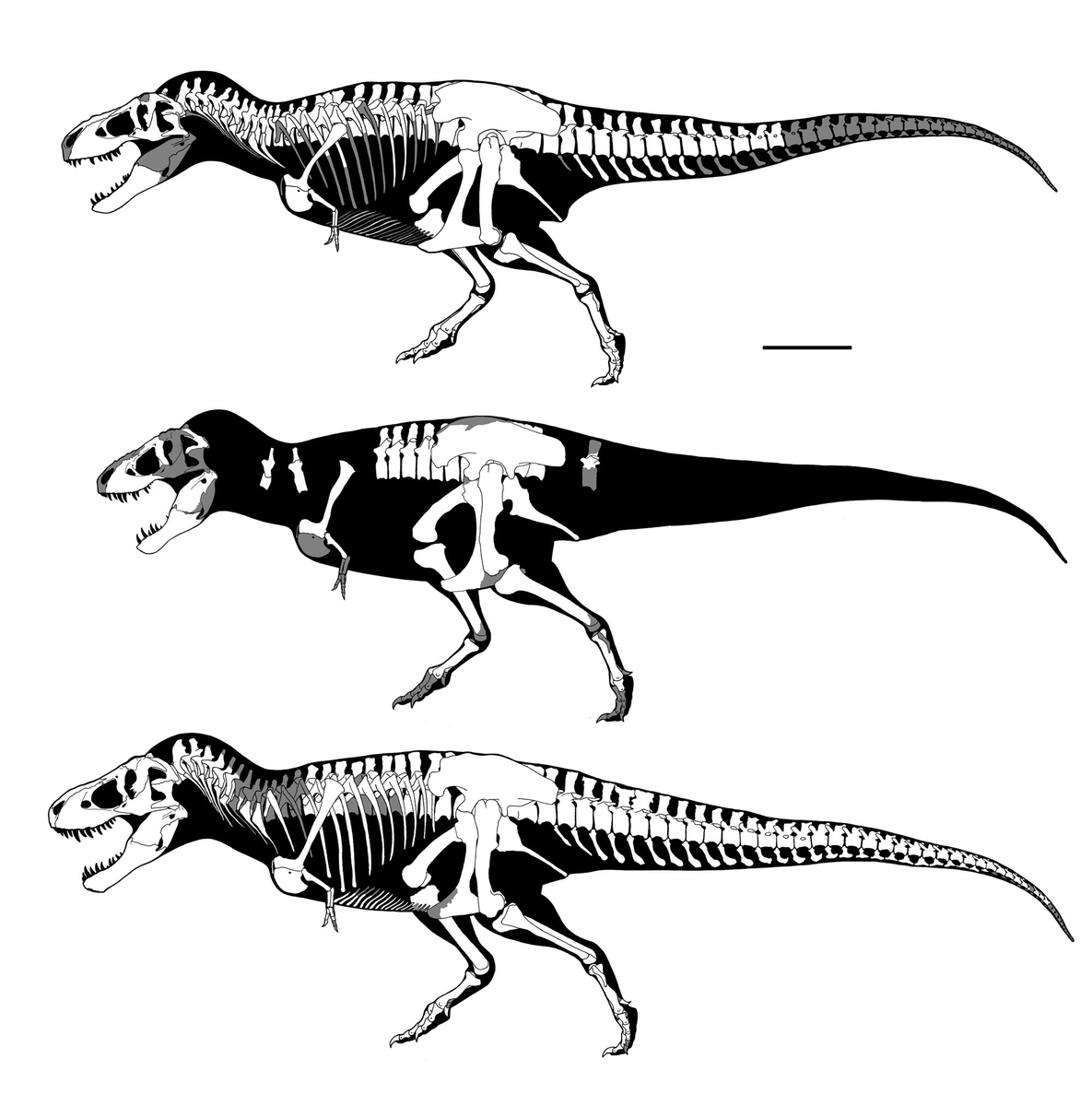
↑Some skulls of "Monoclonius". Scale bars are 1m. AMNH 5239 is the holotype of Monoclonius flexus.
角竜の研究が始まって150年が過ぎようとしているが、その中にあって、初期の研究の産物を代表するものがモノクロニウスである。筆者くらいまでの世代であれば(エウセントロサウルスとセットで)子供向けの恐竜図鑑でもギリギリ見たことがあるだろう。
不穏な記事タイトルから察していただけようが、恐竜図鑑から消えて久しい昨今にあって、モノクロニウスはなおも怪しく蠢いている。悪夢のはじまりを告げたこの恐竜の再整理が済んではじめて、ジュディス・リバーの悪夢を振り払ったことになる――が、その日はまだ見えない。
第一次化石戦争真っ只中の1876年、まだ20代のチャールズ・ヘイゼリアス・スターンバーグとジョン・C・イザーク(ほとんど無名である)を伴い、コープはモンタナ州ジュディス・リバーの近辺で調査を行っていた。あちこちを巡り歩く間にそれなりの量の化石――恐竜については寄せ集めの歯が多かったが、植物食恐竜の骨格要素もそれなりにあった――が集まり、同年のうちにコープはこれらを記載することにした。アウブリソドン、ラエラプス(この時点ではまだトゲダニ属のホモニムだとは発覚していなかった)、ディスガヌス(新属)と不穏な名前が続く中(いずれも歯に基づく記載である)、いくらかの歯と寄せ集めの骨格要素に基づきモノクロニウス・クラッススMonoclonius crassusを命名したのであった。
コープはいつものごとく、やたら簡潔(と見せかけてその実意味不明だったりもするのだが;後述)な論文(図は1枚もない)をもってモノクロニウスを記載した。歯の形態はハドロサウルスによく似ているが、一方で頑丈な前肢と長大な腸骨はハドロサウルスのそれとはまったく異なっていた。モノクロニウスの化石にはほかに仙椎、3個の癒合した「前方胴椎」、そして奇妙な「胸骨柄」が含まれており、すらりとした二足歩行のハドロサウルスとはずいぶん異なった見てくれであるように思われた。
(コープが記載したこれらの標本「群」――のちのAMNH 3998には、実のところ鼻角や上眼窩角の要素は全く含まれていない。学名の由来について地の文でそれとなくほのめかすのがコープの記載スタイルであり、何をもってモノクロニウスMonoclonius――「一つの芽」を命名したのか、確実なことはわからない。とはいえ、コープは同じ論文の中でモノクロニウスと対になる属名――「二つの芽」――ディクロニウスDicloniusを歯に基づいて命名しており、モノクロニウスが歯の形態にちなんだ属名であることは確かなようだ。どうもコープは、歯の交換様式に基づいてそれぞれを命名したらしい。コープがこの標本群の中で最も重要視していたのが歯であったともいえるわけだが、読者の方はすでにお気づきの通り、この歯――寄せ集めの標本群の中で事実上の模式標本として機能するはずだった――はハドロサウルス類のものであった。なお、1860年にはライディが角竜の歯をトラコドン・ミラビリスに含めるやらかしをしていたりもする。コープが胸骨関連の要素とみなしたものこそ紛れもないセントロサウルス類のフリルであったが、この時点では側方バーが接合されておらず、奇妙なリッジのあるY字型の馬鹿でかい骨といった様相であった。胸部の要素だと誤認するのもわからなくはない。)
翌1877年、ハイデンによって取りまとめられた先住民居留区の地質・地理に関する大著の中で、コープは初めてモノクロニウスの図示を行った――が、これは前年に記載したものとは別の標本(のちのAMNH 3999)であった。この標本(これは寄せ集めではなく、まとまった単一個体の要素である)について、脳函と眼窩まわり(小さめの上眼窩角を含む)の図示を行ったコープであったが、(脳函について、鳥類との類似を指摘しつつも)分類を確定することができなかった。歯はディクロニウスに、図示しなかった首から後ろの要素はモノクロニウスやアガタウマスに似ていたが、コープは(なぜか)ここで新属を立てたりすることはなかったのである。コープは上眼窩角の存在を認識しつつ(角芯であるかさえ自信がなかった)、それが頭骨のどの部位にあたるのかさえ決めかねたのであった。角竜の実態にかなり迫ってはいたのだが、珍しく慎重だったコープはマーシュを出し抜く千載一遇のチャンスを逸したのである。
(ケラトプス科角竜といえば頭骨ばかり見つかるイメージがあるが、概形のわかる「まとも」な頭骨の発見は1889年の“ケラトプス”・ホリドゥスを待たねばならなかった。ここからハッチャーによる怒涛の発見が続くわけで、要は注目している産地の問題だったのである。150年以上にわたる研究の歴史にあって、ジュディス・リバーで発見された1個体分のまともな角竜の頭骨はわずか数点である。)
それからの数年でディクロニウスが巨大なハドロサウルス類であるらしいことが判明した一方、モノクロニウスの正体は判然としなかった。1886年になってコープは“ディクロニウス・ミラビリス”(AMNH 5730:のちのアナトティタン・コープイのホロタイプにして現エドモントサウルス・アネクテンス)の胸骨と共に、初めてモノクロニウス・クラッススの「胸骨」を図示したが、依然として「ハドロサウルスに似た歯を持つ」恐竜であると考えていたのである。
状況が変わったのは1888年であった。マーシュの右腕としてジュディス・リバーへ乗り込んだジョン・ベル・ハッチャーが、一対の角(破損して1本ずつに分かれていたが)のある頭蓋天井と、それに続く後頭顆を発見したのである。1888年の暮れ、マーシュはこれを新属新種ケラトプス・モンタヌスCeratops montanusとして記載・図示を行い、またステゴサウルスに近い新たなグループ――ケラトプス科を設立した。ここに初めて、角のある植物食恐竜の存在が認識されたのである。
(マーシュ自身、ケラトプスが初めて記載する角竜だったわけではない。この前年、ジョージ・キャノンJr.によってコロラドはデンヴァーにて採集された見事な一対の角――ケラトプスとは違って頭蓋天井とつながっていた――を、鮮新世後期のバイソン属の新種、ビソン・アルティコルニスBison alticornisとして記載していたのである。トリケラトプス属を設立する際にマーシュはこの同定を訂正したが、なぜかトリケラトプスではなくケラトプス属として再分類した。このあたりの下りは前記事を参照)
ケラトプスの記載を見るなり、コープは1877年に図示した化石の正体に気が付いた。慌てたコープは即座に短報を出し、この標本がケラトプスに近縁であること、そして1874年に命名していたポリオナクス・モルトゥアリウスPolyonax mortuariusもこの類である可能性を述べたのであった。
(コープはこの時点でも、1877年に図示した標本を命名することはなかった。コープは断片の寄せ集めたるポリオナクスに角の断片らしいものがあることに触れている(ハッチャーらもこれに追随した)が、結局のところこれは腓骨の破片であるようだ。)
年が明けた1889年、絶好調のハッチャーはついに角竜の「まとも」な頭骨の発見に至った。ワイオミングの“ララミー層”で発見されたそれは、だいぶ破片化していたものの、2本の巨大な上眼窩角と、1本の鼻角――3本の角を保存していたのである。このケラトプス属の新種――ケラトプス・ホリドゥスを皮切りに、ララミー層では3本の角を備えた頭骨が次々と見つかるようになった。夏になるとマーシュがケラトプス・ホリドゥスのために新属――トリケラトプスを設け、12月にはついにトリケラトプス属の頭骨を完全に復元できるようになった(ここで初めてトリケラトプス属――トリケラトプス・“フラベラトゥス”の図示が行われた)。3本の角、鋭いくちばし、力強い顎、そして巨大なフリル――角竜の姿が明らかになった瞬間であった。
ここに至り、ようやくコープも、モノクロニウスの「胸骨」がフリルであることを認識した。1889年の8月付の論文(実際に出版されたのは1890年であるし、執筆自体も1889年12月のマーシュの論文を読んでからである)にて、コープはモノクロニウス・クラッススの再記載を行うとともに、モノクロニウス属の新種の記載・図示を行った。1877年に記載・図示した標本――モノクロニウス・レクルヴィコルニスMonoclonius recurvicornisは脳函と上眼窩角に加えて鼻角(斜め上方へ伸び、そこから下方へカーブしている)と吻の一部を保存しており、またモノクロニウス・スフェノケルスM. sphenocerusは見事な鼻角と吻の一部をよく残していた。モノクロニウス・フィススM. fisusはだいぶ小さな個体であり、鱗状骨を保存しているかに見えた(が、これは翼状骨の誤認であった)。コープはさらに、1872年に首なしの骨格に基づき命名していたアガタウマス・シルヴェストリスが、モノクロニウス・クラッススの椎骨とよく似ていることを指摘し、これもモノクロニウスやポリオナクスの近縁とみなした。そしてコープはこれら角竜を1888年に命名されたばかりのケラトプスではなく、そのうちでもっとも古くに命名された属――アガタウマスに基づく名前である、アガタウマス科と呼ぶべきであると主張した。
(コープはこの時、M.スフェノケルスをモノクロニウス属ではなくアガタウマス属に分類する可能性について触れている。このことが巡り巡ってナイトによるアガタウマスの復元画――トリケラトプス・プロルススのフリルとM.スフェノケルスの鼻角を組み合わせたもの――へつながっていく。)
続いてコープは(論文は1889年10月付ではあるが、執筆は1890年の3月であった)ポリオナクスがトリケラトプスのシニアシノニムであることを主張し、またモノクロニウスをケラトプスのシニアシノニムとした。コープいうところのアガタウマス科の構成メンバーはかくして、長い上眼窩角と短い鼻角、窓のないフリルを持つポリオナクス、短い上眼窩角と長い鼻角、窓の開いたフリルを持つモノクロニウス、そして巨大なアガタウマスとなった。マーシュの命名による分類群をすっかり片付け、コープはようやく留飲を下げたのだった。
このあたりのコープの動きは当然マーシュには無視された。ポリオナクス・モルトゥアリウスの模式標本は破片を通り越して残骸もいいところであり、モノクロニウスにしてもM.レクルヴィコルニスはケラトプス・モンタヌスと比較可能な部位は眼窩まわりだけ(後述)に過ぎなかったのである。1895年になってコープのコレクションのかなりの数がAMNHに売却され、ハッチャーによって再検討が行われることとなった。
コープ自身認めていたことではあったのだが、AMNH 3998――モノクロニウス・クラッススの原記載に用いられた標本は、明らかに複数個体(同じ場所で産出したかどうかすら怪しい)の寄せ集めであった(さらに言えば、コープが種々の記載で用いた標本が具体的にどれなのか確認することさえ難しい状況でもあった。コープの記載には図はおろか、標本番号の指定すらなかったのである)。とりあえず歯以外の要素は角竜で間違いないようだったが、首から後ろのばらけた要素でケラトプス科の属種を決定することが困難であることはハッチャーにはわかりきったことであった。
苦闘の末、コープが記載に用いたらしい標本をいくらか見つけ出したハッチャーにより、ようやくモノクロニウスのまともな記載が行われることになった(出版の前にハッチャーは腸チフスで無惨に死んだ)。ハッチャーはAMNH 3998――モノクロニウスのシンタイプ(複数個体の混じった基準標本)からフリルのみをレクトタイプとして選び出し、将来の大混乱への備えとした(コープがM.クラッススの上眼窩角とみなしていたらしい標本AMNH 3997は、明らかにフリルの主よりも小さな個体のものであった)。また、M.レクルヴィコルニスをとりあえずケラトプス属へ移動させることも提案している。
さて、これに先んじる1902年、ランベによってカナダからモノクロニウス属の新種が3種報告されていた。モノクロニウス・ドーソニM. dawsoniは吻を欠く以外はほぼ完全な頭骨(フリルは風化して崩れかけており、採集することはかなわなかった――これが新たな火種となった)CMN 1173に基づいており、またほぼ完全なフリルと鼻角らしき断片からなるCMN 971もM. ドーソニとみなされた。このフリルはM. クラッススのそれと比べてずっと派手な装飾が特徴であった。モノクロニウス・ベリM. belliはT字型のフリルの断片CMN 491に基づいており、M. クラッススと比べてずっと長いフリルを持っていたらしかった。モノクロニウス・カナデンシスM. canadensis(CMN 1254)は吻と頭頂骨の大半を欠いていたが、短いフリルと短くそして後方へカーブした上眼窩角を備えていた。ハッチャー(とラル)はこれらについても再検討を加え、モノクロニウス・ベリがトロサウルス属に似たフリルを持っていたらしいこと、モノクロニウス・ベリとモノクロニウス・カナデンシスをケラトプス属に移動すべきことを述べたのだった。
これと前後する1904年、ランベはCMN 971をモノクロニウス・ドーソニから切り離し、新属新種セントロサウルス・アペルトゥスとして記載した(上記の項を執筆中だったハッチャーのアドバイスがあったらしい)。ラルはこれを受け入れ、モノクロニウスとは明確に異なるものとしてセントロサウルスを扱うこととしたのである。
ハッチャーの遺稿はラルによって仕上げられ、1907年にひとまずの角竜の総まとめが出版された。第一次化石戦争の終結から10年余りを経て、いちおうの戦後処理が終わった格好であった。
が、一息付けたのはここまでであった。1912年にはブラウン率いるAMNH隊が本格的にアルバータ入りし、次いで60代になっていたチャールズ・ヘイゼリアス率いるスターンバーグ一家もブラウン隊を追いかけてレッド・ディアー川へと漕ぎ出したのである。第二次化石戦争が始まったのだった。
(1910年になり、ランベはセントロサウルス・アペルトゥスの復元を訂正している。CMN 971の鼻角とされていた「角」は、実際にはフリルから垂れ下がるように長く伸びたホーンレット(P1)であった。1909年の夏にランベを訪ねたブラウンが、頭頂骨に新鮮な破断面を発見し、「鼻角」がそこにぴったりはまることに気付いたのである。)
モンタナとは違い、アルバータの「ジュディス・リバー層」(のちベリー・リバー層、現ダイナソー・パーク層)では完全な角竜の頭骨がいくらでも見つかった。関節のつながったほぼ完全な骨格さえ見つかり、ケラトプス科角竜の解剖学的特徴に関する理解はほんの数年で飛躍的に高まったのである。
ブラウンらは1914年に見事な角竜の頭骨AMNH 5239を記載するにあたり、まずモノクロニウス・ドーソニとセントロサウルス・アペルトゥスが結局は同じ種に属するらしいことを指摘した。セントロサウルス・アペルトゥスを(もともとランベが考えていたように)モノクロニウス・ドーソニのシノニムとしたうえで、AMNH 5239――セントロサウルス・アペルトゥスに酷似したフリルを備えていた――を新種モノクロニウス・フレクススM. flexusとして記載したのである。モノクロニウス・クラッススのP1やP2は風化かなにかで死後失われたとみなし、ランベがセントロサウルス属をモノクロニウスから分割するのに用いたはずの特徴をモノクロニウス属本来の特徴として扱ったのであった。
これと前後して、ランベは自ら命名したモノクロニウス属の種の整理を進めていた。ハッチャーらの指摘した通りM. ベリはより新しい時代のトロサウルスに続く系譜にあるように思われ、ランベはこれにプロトロサウルスProtorosaurusの属名を与えた――が、これはペルム紀の主竜形類にとっくに用いられており、すぐにカスモサウルスChasmosaurusの属名をM. ベリに与えた。ランベはまた、M. カナデンシスがトリケラトプスと似たフリルを持っていることを指摘し、これをトリケラトプスの祖先とみなして新たな属名――エオケラトプスEoceratopsの属名を与えたのだった。
ランベは自らの仕事を片付ける中で、極めて重要な指摘――ケラトプスとモノクロニウスの模式種がそれぞれ貧弱な化石に基づいていることを述べている。ケラトプス・モンタヌスはそっくりフリルを欠いており、モノクロニウス・クラッススは(ハッチャーの苦闘を経ても、依然として)標本に付随してくる情報がほとんどなかった。ランベはこれらの属名を用いるべきではないこと――疑問名として扱うべきことを提案したのである(ランベの分類の大枠はハッチャーらの影響を強く受けていたが、ゆえにモノクロニウス・カナデンシスをケラトプス・カナデンシスとするのではなく、新属エオケラトプスを設けた)。ランベはモノクロニウス・ドーソニをブラキケラトプス属へと再分類し、セントロサウルス・アペルトゥスと明確に区別したのであった。ランベはまた、前方に曲がった鼻角と短いが明瞭な上眼窩角を持った頭骨をセントロサウルス・アペルトゥスとして記載し、M.フレクススがC.アペルトゥスのシノニムに過ぎないことを示した。
ブラウンはこれを受け入れることなく、1917年になってほぼ完全かつ関節のよくつながった標本AMNH 5351をモノクロニウス・ナシコルヌスM. nasicornus、首なしのミイラ化石AMNH 5427をモノクロニウス・カトラーリM. cutleriとして記載した。M. フレクススはその種小名の通り前方へ折れ曲がった鼻角を持つ一方、M. ナシコルヌスはM.スフェノケルスとよく似た真っすぐな鼻角を備えていた。そしてM. ナシコルヌスのフリルは“セントロサウルス・アペルトゥス”やM. フレクススと同様の形態――垂れ下がったP1と、内側向きにまがったP2を備えていた
そんなわけでセントロサウルスをモノクロニウスのシノニムとした(というよりは、セントロサウルス属の特徴とされていたものをそのままモノクロニウス属にスライドしただけの)ブラウンと、モノクロニウスを疑問名としてセントロサウルスを残したランベとで分類のコンセプトはきっぱり分かれることとなった。この状況はだいぶ尾を引き、1933年になってラルが出版した新たな角竜のモノグラフ(1907年の増補版をうたっていたが、こちらは名実ともにラル一人の仕事である)では、モノクロニウス属の下にセントロサウルス亜属を設ける始末であった。ラルはコープが命名した一連のジュディス・リバー産の種(とM.ドーソニ)を全て(それぞれ独立種としたうえで)モノクロニウス亜属とし、ベリー・リバー産の種(ブラウンがモノクロニウス属として命名したものを含む)を(これも全て独立種としたうえで)セントロサウルス亜属としたのである。
第二次化石戦争の火もすっかり消えた1930年代後半にあって、チャールズ・モートラムはまだレッド・ディアー川流域を掘り続けていたが、ここで巨大なモノクロニウスらしき頭骨に出くわした。この頭骨CMN 8790はモノクロニウス・クラッススと酷似したフリルを備えていたが、侵食によってP1を失った形跡はなかったのだった。チャールズ・モートラムはCMN 8790をモノクロニウス・ロウイM. loweiとして記載し、モノクロニウスとセントロサウルスが明確に異なることを指摘した。彼はまたセントロサウルス属の新種セントロサウルス・ロンギロストリスC. longirostrisを設け、新標本に基づきM.ドーソニをセントロサウルス属へと移した。こうして状況は落ち着いたかに見えた。
デジタル的な手法――分岐分析がその最たるものである――が古生物学に導入された1970年代以降、様々な視点から古典的な分類群の再整理が進んだ。ボーンベッドから産出した標本の検討が行われる中で、ケラトプス科角竜は個体変異が著しいらしいことが示されるようになり、属種の整理が始まったのである。
1990年になり、ドッドソンが分岐分析の手法で取り組んだのがモノクロニウスとセントロサウルスの再整理であった。ドッドソンはモノクロニウス・レクルヴィコルニスとM.フィスス、それからセントロサウルス・カトラーリ――頭骨が著しく不完全か、あるいは首なしだった――を疑問名とし、それからM. ロウイをモノクロニウス・クラッススのシノニムとした(これでモノクロニウス・クラッススの完全な頭骨が知られている格好になる;M. スフェノケルスは?付きでM. クラッススのシノニムとした)。セントロサウルス属はC. アペルトゥス一種にまとめつつ、C. ナシコルヌスはスティラコサウルス・アルバーテンシスのメスとみなした。
(この当時、様々な恐竜の雌雄をなんとかして識別しようという研究がひたすらに流行っていた。C.ナシコルヌスをスティラコサウルス・アルバーテンシスとするドッドソンの意見はその後全く顧みられず、またこの当時様々な研究者によってなされた角竜の雌雄差診断も肯定的には見られていない。)
なんにせよドッドソンのこの研究で、モノクロニウスとセントロサウルスの違いが再確認されることになった。ついでに両者とも1属1種となり、ずいぶんすっきりしたわけである。
一方、これと前後して、ランベが1902年に設けたセントロサウルスCentrosaurusの属名が、1843年には既に別の動物に対して用いられていたらしいことが明らかになっていた。両生類・爬虫類の分類の大家であったレオポルト・フィッツジンガーが、テキサスツノトカゲPhrynosoma cornutumのジュニアシノニムとして、セントロサウルス・ホリドゥスCentrosaurus horridusを使っていたのである。かくしてチューレとマッキントッシュは、角竜としてのセントロサウルスの属名をエウセントロサウルスEucentrosaurusの新属名に置き換えることを提案した(ヘロプスに対するエウヘロプスしかり、よく用いられる手ではある)。
しかしこれはいまいち定着せず、文献によってセントロサウルスとエウセントロサウルスの属名が入り乱れる結果となった。セントロサウルス・ホリドゥスの名は1843年以前に(=プリノソマ・コルナトゥムのジュニアシノニムになる前に)使用された形跡がなかったのである。フィッツジンガーはどういうわけか(少なくとも出版された限りでは)、わざわざ初めからP. コルナトゥムのジュニアシノニムとしてセントロサウルス・ホリドゥスの名を設けたらしかった。フィッツジンガーのセントロサウルス属が分類名として機能したことは事実上一度もなかったことが確認され、結局はランベのセントロサウルス属が有効性を保つこととなったのである。結果、エウセントロサウルスの属名はその存在意義を失い、2000年を過ぎるころには単にランベのセントロサウルス属のジュニアシノニムとして扱われるようになった。
さて、90年代になってエイニオサウルスとアケロウサウルスが発見され、またセントロサウルスのボーンベッドも新たに発見されるようになると(パイプストーン・クリークの話も言わずもがな)、角竜の成長に関する研究に火が付いた。ボーンベッドからは様々な成長段階のセントロサウルス類の化石が採集されたが、未成熟個体らしいもののフリルはことごとくブラキケラトプスやモノクロニウスと酷似していたのである。ここに至り、ブラキケラトプスはおろかモノクロニウスまでもがなんらかのセントロサウルス類――なんらかの新種かもしれないし、既知のものかもしれないが、それさえ判断できそうもない――の幼体と亜成体であるらしいことが明確に示されたのであった。
ドッドソンがこれをすんなり受け入れることはなく、モノクロニウス(この場合M.ロウイを意味する)がセントロサウルス類の亜成体にしてはむやみに大きいことを指摘した。モノクロニウスの成体と目される他の角竜と比べても大きかったのである。ドッドソンはモノクロニウスの形態が幼形成熟の産物であり、様々なセントロサウルス類のボーンベッドから産出した「モノクロニウス様」の亜成体とは別物――独立属であると主張したが、とはいえ、もはやモノクロニウス・クラッススとM. ロウイを積極的に結びつける証拠は何もなかった。モノクロニウス・クラッススのホロタイプは特別巨大なわけでもなかったし、モノクロニウス・ロウイも巨大なだけで明らかに未成熟――頭骨の癒合がほとんど進んでいなかったのである。The Dinosauria第2版――2004年刊行――ではモノクロニウス・クラッススは「おそらく有効名」として扱われたが、それがモノクロニウスの最期であった。命名から130年を経て、ついにモノクロニウス――事実上角竜に基づいた属名ではなかった――は、疑問名としてコンセンサスを得たのである。ランベによる最初の提案から、90年の歳月が流れていた。
かくして今日、モノクロニウスは疑問名として扱われている。そうはいってもあくまで学名の取り扱いの話であり、セントロサウルスのシノニムになりそこなったモノクロニウス属の標本は今日もそこにあり続けている。
年代測定の手法の拡大や高精度化に伴い、特に北米においては恐竜の分類学的再検討について生層序学的な観点も加えられるようになった。コープの命名した一連のモノクロニウス属はジュディス・リバーJudith River層のコール・リッジCoal Ridge部層下部(ざっと7600万年前ごろ)から産出した可能性が高い(現状で産地の再特定には至っていないのだが)のだが、これはダイナソー・パーク層では最上部――スティラコサウルス・アルバーテンシスが姿を消し、突如(先の記事で述べた)パキリノサウルス類が姿を現す(そして直後に海成層であるベアポウBearpaw層が堆積する)――に対比される層準でもある。“モノクロニウス・ロウイ”も、ちょうどこのあたりの層準からの産出である(むしろパキリノサウルス類よりも新しい可能性もある)。ジュディス・リバー層より内陸であるトゥー・メディスン層にはこの時期“スティラコサウルス・オヴァトゥス”(結局のところスティラコサウルス・アルバーテンシスに過ぎないかもしれない)や“ブラキケラトプス”、そしてステラサウルスあたりがいたかどうかといったところであり、“モノクロニウス”――単一種とは限らない――の成体候補はすんなり見つからない。
“モノクロニウス・ロウイ”は吻が長く、(セントロサウルスの亜成体よりも)スティラコサウルスの亜成体によく似ている。P3だけが長く伸びそうな気配を見せているあたりはスティラコサウルスの亜成体と異なる(サイズもこちらの方が大きい)が、時代からしても、スティラコサウルスやその系譜――ステラサウルスやエイニオサウルスではP3が長く伸びる――に近いのかもしれない。一方で、“モノクロニウス・スフェノケルス”はよりセントロサウルス的な吻の長さのようである。つい最近になってジュディス・リバー層(層準不祥)で紛れもないセントロサウルスが産出したというのだが、あるいはこれとの関係があるのかもしれない。“モノクロニウス・レクルヴィコルニス”でみられる、短いが明確な上眼窩角と前傾した鼻角の組み合わせはまごうことなきセントロサウルス・アペルトゥスで知られている。“モノクロニウス・クラッスス”のフリルの形態は派生的なセントロサウルス類の亜成体のそれであり、なにかしらの独自性を見出すのはやはり困難なようだ。
“モノクロニウス”たちの真の姿を明らかにするうえで、産出層準の特定と、それと対比される層準での成体探しは避けては通れない。そして、“モノクロニウス”たちの産出層準が明らかになり、かつそれと対比される層準でセントロサウルス類の成体の完全な頭骨が発見されたところで、結局は“モノクロニウス”――セントロサウルス類の亜成体や断片の寄せ集め――との比較は非常に難しいのである(だからこそ疑問名というシステムが存在する)。
“モノクロニウス”たちが疑問名を脱し、分類学的な議論の俎上に再び上がることは、おそらくこの先もないだろう。それでもなお、ケラトプス科角竜の分類に携わる研究者たちは、ジュディス・リバー層――ケラトプス科のふるさとで、醒めない悪夢を見続けている。
(“モノクロニウス・ロウイ”はセントロサウルス類としては驚くほど長いフリルを持っており、しばしば本種の独自性らしいものとして挙げられる――が、見まごうことなきセントロサウルス・アペルトゥスの亜成体で長めのフリルを持っている標本が知られており、また最近記載されたエイニオサウルスないしアケロウサウルスの亜成体と思しき標本も“モノクロニウス・ロウイ”と互角以上の長いフリルを持っている。成長のある時期にフリルの伸長が先んじるというだけのことのようだ。)




{kind=link}