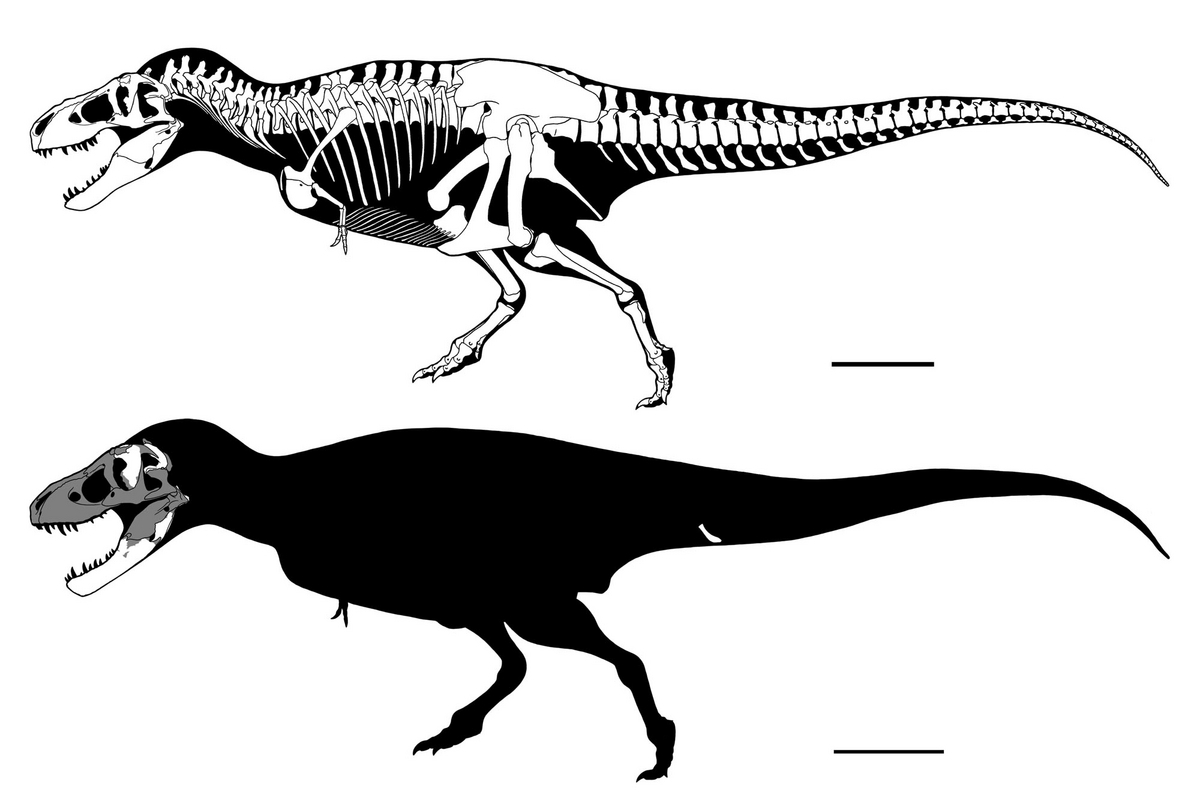
↑ティラノサウルス・レックスTyrannosaurus rexのホロタイプCM 9380(旧AMNH 973)の骨格図。マッシブな体型に注意。
命名120周年である。今日も今日とて人気を集めているティラノサウルス・レックスだが、命名当初はさほど有名だったわけではない――というありがちなイントロは、本種にはまったく当てはまらない。稀代のプロデューサーによって舞台をあつらえられたそれは、命名されるずっと以前から研究者たちに待ち望まれていた存在でさえあったのである。そして数奇な運命をたどったホロタイプCM 9380は、「最初のティラノサウルス・レックス」としてあるべき役割を今日も果たし続けているのだ。
1854年から1855年にかけ、「転がる石を拾う男」ことフェルディナンド・ヴァンディヴィア・ヘイデンはネブラスカ準州はジュディス川流域のバッドランドで地質調査(時代が時代なので事実上の「探検」である)中であった。ジュディス・リバーJudith River層を踏査するうち、ヘイデンはそれなりの数の脊椎動物の小さな化石を手にしていた。一連の化石はペンシルバニア大学の若き解剖学教授であったジョセフ・メリック・ライディの下へと送られ、ライディは1856年にこれらを8つの種に分類した。この中には北米で初めてとなる恐竜化石(いずれも遊離歯)が4種含まれていた――パラエオスキンクス・コスタトゥスPalaeoscincus costatus、トラコドン・ミラビリスTrachodon mirabilis、トロオドン・フォルモススTroodon formosusそしてデイノドン・ホリドゥスDeinodon horridusである。
(ライディによる1856年の論文は2ページの簡潔な記載に留まっており、標本の図示もない。1860年になって初めてこれらの標本の図示が行われたのである。ライディはパラエオスキンクスを全長30フィート以上の超巨大な半水生のイグアナ科、トロオドンを大型の半水生のオオトカゲとみていた。ヘイデンの採集したこれらの化石、およびその後コープのチームが採集した一連のジュディス・リバー層産の恐竜化石は、すべてマクレラン・フェリーMcClelland Ferry部層上部からコール・リッジCoal Ridge部層下部にかけての層準から産出したものとみられている。これらの部層の境界(“ジュディス中部不連続面Mid-Judith Discontinuity”)はカナダ・アルバータ州は州立恐竜公園におけるオールドマンOldman層とダイナソー・パークDinosaur Park層の境界と一致すると考えられており、その年代は約7650万~7630万年前ごろとみられている。)
ライディはトラコドンの歯がイグアノドンに、デイノドンの歯がメガロサウルスに似ていることを正確に見て取っていた。1857年にデイノドンをメガロサウルスの亜属に下げるなどの気の迷いもあったが、どうあれこれらのデイノドンの歯はメガロサウルスに似た大型の肉食恐竜が白亜紀のアメリカ西部に存在したことをはっきりと示していた。
が、デイノドンのまとまった骨格は待てど暮らせど西部では出てこなかった。1866年にはニュージャージーはニュー・エジプトNew Egypt層(マーストリヒチアン後期;約6950万~6604万年前ごろ)の最上部でラエラプス・アクイルングイスLaelaps aquilunguisの(ひどく不完全だが)顎と四肢を含むまとまった骨格が産出する一方で、コープ自身やスターンバーグをはじめとするコープ麾下の化石ハンターたちが繰り出していったにもかかわらず、西部では1870年代の中ごろになってもラエラプスの歯がせいぜいだったのである。
(デイノドンのもともとのシンタイプ(ANSP 9530-9544)はティラノサウルス科の上顎骨歯/歯骨歯と(D字/U字断面の)前上顎骨歯が混在しており、同一個体(あるいは種)に由来するものでもない。ティラノサウルス上科の歯と顎がセットで産出したのはラエラプス・アクイルングイス=ドリプトサウルス・アクイルングイスのホロタイプが初めてのケースであったが、これには前上顎骨歯が含まれておらず、コープはゆえにラエラプスがデイノドンとは異なる分類群だと判断した格好である(言うまでもなく、当時ティラノサウルス上科の歯の全容は分かっていなかった)。一方でライディは1868年になってANSP 9533-9535(いずれも前上顎骨歯;9533と9534には鋸歯がある)をデイノドン・ホリドゥスから切り離し、アウブリソドン・ミランドゥスと命名した。これによってデイノドンの模式標本はラエラプスによく似た上顎骨歯/歯骨歯だけになった――が、コープは師のこの分類に噛みつき、ラエラプスの記載の際にデイノドンの「典型」をANSP 9533-9535としたことを指摘した(それ以外の、つまりライディがデイノドンに残留させた要素がメガロサウルスそしてラエラプスと酷似していることはライディ自身認めるところであった)。つまり、コープの弁ではアウブリソドン・ミランドゥスはデイノドン・ホリドゥスの客観的同物異名ということになり、ライディ言うところのデイノドン残留組はすべてラエラプス属ということになる。コープはさらにデイノドンDeinodonがディノドンDinodon(旧マダラヘビ属)のジュニアシノニムであるとまで主張し始め(コープはどういうわけかデイノドンを常にDinodonと綴っていた)、デイノドン・ホリドゥスをアウブリソドン・ホリドゥス――アウブリソドン属はラエラプス属より2年遅く命名されており、よって何があってもラエラプス属が北米産の何かのシノニムになることはない――と呼んだ。ラエラプスの属名にかけるコープのこうした生臭い(しかしあまりにもピュアな)野望はマーシュによってあっさりひっくり返され、デイノドンとアウブリソドンは(実態はさておき)別物として今日扱われている。今日、アウブリソドン・ミランドゥスの扱いについては(言うまでもなくコープの意見を完全無視した)マーシュによるもの――鋸歯を欠いた前上顎骨歯ANSP 9535(比較的最近になり、FMNHに書留郵便で送られる途中で行方不明になった)のみをアウブリソドン・ミランドゥスのレクトタイプとする――が受け入れられている。デイノドン・ホリドゥスはやや宙に浮いた格好となっているが、ともあれ今日ではデイノドン、アウブリソドンのいずれの種も疑問名とされている。)
とはいえ、1870年代の後半になるとコープにも運が向いてきたようだった。コープは1876年に一連のジュディス・リバー層産の新標本をラエラプスの新種として多数命名したが、その中にはラエラプス・インクラッサトゥスLaelaps incrassatus――ザイフォドントziphodontではなくパキドントpachydont的な分厚い、まぎれもないティラノサウルス科の成体の上顎骨歯/歯骨歯が含まれていたのである。さらに同年(実際に出版されたのは翌年であるらしい)には、ジュディス・リバー層からラエラプスの歯と左歯骨のセットを見出した。コープはこの標本AMNH 3963がちょっとした異歯性を持っていることを見抜いていたが、ここに至って不意に思慮深さを発揮した。この標本をラエラプスの新種にするようなことは(不幸にして)せず、単にラエラプス・インクラッサトゥスと同定したのである。
(コープはAMNH 3963をとうとう図示することはなかった。この標本は今日、もっぱらゴルゴサウルス・リブラトゥスのものとされている。どうあれ、ティラノサウルス科と断定できる歯以外の要素としては初めての発見であった。)
さて、上部白亜系に対するマーシュの動きはこのころ静かなものであった。コープがデイノドン属を事実上ラエラプス属に吸収させ、新種を乱発していたたにもかかわらず、マーシュが似たような動きに出ることはなかった(しかしドリプトサウルス属を設立し、コープの手柄をすべてひっくり返すという殺人技を繰り出してはいた)のである。
1873年にコープはヘイデン率いる合衆国地質調査所(USGS)の調査隊の一員としてコロラドの“大褐炭層”――今日のデンヴァーDenver層(マーストリヒチアン後期/ランシアン~ダニアン/トレホニアン;約6820万~6380万年前ごろ)を踏査しており、このとき採集したハドロサウルス類や角竜の断片(言うまでもなくエドモントサウルスやトリケラトプス、トロサウルスであろう)を翌年にまとめて(新種として)記載した。こうした流れがあったにもかかわらず、日ごろの根回しがあったのか、運はマーシュの方に転がってきた。1873年の暮れ、デンヴァーのとなりのゴールデン在住のエドワード・ベルソーという男から、マーシュに恐竜化石の発見を知らせる手紙が届いたのである。
すでに1867年には一帯で採集した化石をスミソニアンへと送っていたベルソーは、マーシュに「牙」(明らかに角竜の角の骨芯である)をはじめとする多数の化石があることを知らせていた。1874年の6月、ベルソーは新たな化石――ラエラプスに似た巨大な歯のスケッチをマーシュに送った。保存のそれほどよくないこの標本YPM 4192は、しかしまぎれもないティラノサウルス・レックスの、初めて発見された化石であった。
ベルソーはデンヴァー周辺で次々と化石産地を見出したが、マーシュがそこで発見される化石に目を向けるようになるにはもう少し時間がかかった。マーシュがYPM 4192を記載することはなく、一帯で地質調査に励んでいたジョージ・キャノンはベルソーの見出した産地から大型獣脚類の顎(ティラノサウルスで間違いないだろう)を掘り出したりもしていたのだが、これはとうとう記載されることなく行方不明になったのである。
しかし、ベルソーの手紙は究極的には実を結んだ。キャノンとマーシュを、そしてハッチャーとトリケラトプス、そしてティラノサウルスをめぐり合わせることとなったのである。1887年、キャノンはデンヴァーのはずれで一対の巨大な「デンヴァーバイソン」の角を発見したのであった。
ジュディス・リバー層における1888年夏シーズンの結果は、ハッチャーにとっては芳しくないものであったらしい。それなりに興味深い化石はあったが、スターンバーグ(父)をはじめとするコープ麾下の化石ハンターたちの成果を圧倒的に上回るようなものではなかった――が、それでも重要な化石がマーシュの手元に届くことになった。この中にはケラトプス・モンタヌスCeratops montanus――「角竜」として初めて認識されることになった標本USNM 2411が含まれていたのである。
1889年の1月にニューヘイヴンに戻ってきたハッチャーはそこで初めて「デンヴァーバイソン」ことビソン・アルティコルニスBison alticornis YPM 1871E(現USNM 4739)と対面し、これがUSNM 2411――産出層からして明らかに哺乳類のものではなかった――の類縁であること、そしてワイオミングはランスLance層(マーストリヒチアン後期;約6910万~6604万年前ごろ)の中部で地元コレクターのガーンジーによって発見された頭骨――のちのYPM 1820もこれと酷似した角を持つことを見て取った。かくしてハッチャーとそのチームはランス層へと導かれることとなったのである。

↑ティラノサウルス・レックスUSNM 2100(左)とUSNM 6183(右)。スケールバーは1m。
幼い長男を病で亡くしながらもランス層へと突っ込んでいったハッチャーと仲間たち(年下の義兄であるオラフ・ピーターソンも調査に加わっていた)を待っていたのは、大量のトリケラトプスと文字通りの哺乳類の歯の山であった。アリ塚にマイクロサイト由来の化石が(アリによって)集積されていることに気づいたハッチャーは片っ端からアリ塚を崩して回ったが、一方で大きな恐竜化石もそこら中から見出していたのである。
ハッチャーは一連の調査で大型獣脚類の断片的な骨格をいくつか発見していた。1891年に採集されたUSNM 6183は大腿骨長が1mを超える大型のもので、これはドリプトサウルス・アクイルングイスよりはるかに大きく、ドリプトサウルス・インクラッサトゥスよりもさらに大きいように思われた。近辺で発見された腸骨USNM 8064も同様の大型獣脚類(ただしUSNM 6183よりも小さな個体に由来する)のものだったが、1890年に発見された中足骨の断片USNM 2110はこれらをはるかに上回る大型獣脚類のものであるようだった。
マーシュはこれらの標本とよく似たものを1890年にはすでに記載していた。デンヴァー層から産出した手と足に基づくオルニトミムス・ヴェロクスOrnithomimus velox(デンヴァー層産)と第III中足骨の断片に基づくオルニトミムス・テヌイスOrnithomimus tenuisそして巨大な第III中足骨に基づくオルニトミムス・グランディスOrnithomimus grandisである。オルニトミムスの第III中足骨はどれも近位端が鳥のように(マーシュは比較用にシチメンチョウを図示している)第II・第IV中足骨に挟み潰されて後方へと追いやられており、前方からは見えない――アークトメタターサルであった。サイズと形態からして、USNM 2110とUSNM 6183はオルニトミムス・グランディスで間違いないように思われたのである。
マーシュはトリケラトプスそしてトロサウルスと同じ地層――「ケラトプス層」からダントツで世界最大の肉食恐竜の化石が産出したことにいたく感動したようであった。
「この巨大な肉食動物はケラトプス科の天敵のひとつであった可能性が高く、頭骨や骨格全体の発見が興味とともに待たれる。攻撃用に特殊化したそれらの特徴に対抗すべく、ケラトプス科の特異な防御装甲が設計されていることには疑いの余地がない。」
マーシュは予言めいた一節を記したが、とうとう「攻撃用に特殊化したそれらの特徴」――ティラノサウルス・レックスの頭骨を見ないまま、1899年に死んだ。
(オルニトミムス科の頭骨の発見、そして“オルニトミムス”・グランディスがティラノサウルス科(ないしそれにごく近縁なものの寄せ集め)であることが判明したのはだいぶ後――1916年になってからの話である。ランス層産の“オルニトミムス”・グランディスがティラノサウルス・レックスであることはほぼ間違いないが、“オルニトミムス”・テヌイス(少なくともオルニトミムス科ではないようだ)や“オルニトミムス”・グランディスのホロタイプ(スミソニアンに移管されるまでに行方不明になった)のうち、少なくとも後者はジュディス・リバー層ではなくイーグルEagle層(イーグル砂岩とも;サントニアン~カンパニアン前期;約8570万~8120万年前)産であるとみられており、であればリトロナクスLythronaxやディナモテロルDynamoterrorよりも古い、ティラノサウルス科の最古の化石記録の可能性まである。マーシュは実のところ1892年にランス層産の前上顎骨歯に基づきアウブリソドンの新種を2つ命名しており、とりわけ後者はナノティラヌス・ランセンシスであるように思われる――が、これらの形態はそもそもジュディス・リバー層産のものと区別がつかず、そんなわけでナノティラヌス・ランセンシスに優先することはない。)
マーシュによるさりげない必殺の一撃――Laelapsという名が1813年にすでにトゲダニ属として用いられていることを踏まえ、コープの一連のラエラプス属に代わるものとしてドリプトサウルス属を設けた――をコープはどうにか無視し続けた。マーシュがキャリアの絶頂にあった1880年代はコープにとってひたすら耐える時期となり、銀鉱山への投資の失敗やペンシルバニア大との関係悪化もあって経済的に困窮した。しかし1890年代に入ると運はやや上向いてきた(コープの攻撃の甲斐あってマーシュのキャリアにもそれなりに傷がついた)。ハッチャー一行がワイオミングで猛威を振るう中にあって、コープの次の一手はカナダから転がり込んできたのである。
1880年代に行われたカナダ地質調査所(GSC)のティレルとウェストンによるそれぞれの調査によって、今日のカナダはアルバータ州、ホースシュー・キャニオンHoreseshoe Canyon層(当時はララミーLaramie層と呼ばれていた;カンパニアン末~マーストリヒチアン後期;約7310万~6795万年前)で2体のよく似た獣脚類が採集されていた。どちらの標本もぺしゃんこにつぶれていたうえに保存もよくなかったのだが、上下の顎が噛み合ったままの頭骨が残されていた。下顎はどちらの標本も比較的完全であり、ティレルの採集したもの(のちのCMN 5600)では脳函が、ウェストンの採集したもの(CMN 5601)では頭蓋天井を除く頭蓋の大半まで残されていたのである。
当時のカナダは恐竜研究の黎明期にあり、カナダ地質調査所には恐竜化石をまともに扱える専門家がまだいない状況であった。これらの頭骨の記載を要請されたのはコープだったのである。
思いがけず転がり込んできた2つの頭骨を、コープはまたしてもラエラプス・インクラッサトゥスと同定した。頭骨が部分的に含気化していること、(前上顎骨歯は失われていたが)歯骨の先端にある歯がどことなくアウブリソドン(この場合デイノドンのオリジナルのシンタイプの前上顎骨歯すべてを指す)的な「切歯」状であることをしっかりと見抜いたコープであったが、後頭部がまともに残っていなかったこともあり、前眼窩窓を眼窩と誤認した。 「眼窩より前方の領域と長さが等しい」巨大な眼窩の存在について強調したコープは、マーシュが1884年に命名したケラトサウルス・ナシコルニス(中大型獣脚類の保存のよい完全な頭骨が記載されたのはこれが初めてだった)がメガロサウルス属であることを述べ、一連のラエラプス・インクラッサトゥスの頭骨の図示は最終的にカナダ地質調査所の出版物でなされることを記してこの論文を終えたのだった。
結局、コープが「ラエラプスの頭骨」を図示することはとうとうなかった。コープは1896年からたびたび病に伏せるようになったのである。そんな中、親しい教え子であったオズボーンは才気あふれる若きフリーの画家――チャールズ・ナイトをコープに紹介することにした。
コープにとってナイトの復元画の監修は最後の仕事の一つとなった。1897年に完成した「跳躍するラエラプス」――最期までコープはドリプトサウルスとは呼ばなかった――のらんらんと輝く目は、前眼窩窓の中に押し込められていた。
コープの“ラエラプス”に関するやりかけの仕事は、カナダ軍の士官を経てカナダ地質調査所でサイエンスイラストレーターとして名を上げつつあったランベの双肩へとかかってきた。ランベはオズボーンとともにカナダ西部の白亜系層序の整理を進めつつ、これらドリプトサウルス・インクラッサトゥスの頭骨を図示し、コープの記載の誤りを修正し、ティラノサウルス科初となる頭骨の復元図まで1904年に出版することとなったのである。
カナダにおける「化石戦争」のあとしまつはこれくらいで済んだのだが、こうした「戦後処理」は当然アメリカの方がはるかに深刻な問題となっていた。コープにせよマーシュにせよやりかけで終わった仕事はあまりに多く、イェール大学のピーボディ博物館(YPM)およびそこからマーシュのコレクションの相当量を移管されたスミソニアン/国立自然史博物館(USNM)、ライディやコープの初期の研究の核となったフィラデルフィア自然科学アカデミー(ANSP;今日のドレクセル大学自然科学アカデミー)、そしてコープの私蔵コレクションを購入したアメリカ自然史博物館(AMNH)の収蔵庫は混沌とした状態だったのである。
1900年になり、マーシュの後任としてUSGSの古生物学担当を兼任するようになっていたオズボーンは、マーシュの助手からプリンストン大学を経てカーネギー博物館(CM)へと移っていたハッチャーに尻ぬぐい要員としての白羽の矢を立てた。カーネギー博物館ののちの館長であるホランドとは激しい学術的・政治的対立のもとにあったオズボーンだが、ハッチャーやそのかつてのチームの一員たちを高く評価していたのである(AMNHの古脊椎動物学部門の立ち上げ時にハッチャーの引き抜きを試みたことさえあった)。ハッチャーはマーシュのほぼ手付かずだった仕事――USGSが出版を予定していた角竜のモノグラフの執筆を快諾し、コープの記載した角竜の再記載のためにAMNHを訪れるようになったのだった。
コープの論文は図版に乏しいことで当時から悪名高く(マーシュの論文にしても、今日からしてみれば驚くほど図版に乏しいのだが)、ハッチャーは読みやすいとはいえない記載を頼りにAMNHの収蔵庫をさまよう日々が続いた。そんな中でハッチャーが目を留めたのは、マノスポンディルス・ギガスManospondylus gigas――コープが1892年にアガタウマス科(ケラトプス科と実質的に同義)の新属新種として記載した椎骨AMNH 3982であった。ハッチャーが採集しマーシュが記載してきたどんな角竜よりも巨大なものとして記載されていたそれは、しかしどこからどう見ても獣脚類の椎骨であった。2個あるはずだった椎骨はいつの間にかひとつだけになっていたのだが、破損した面から見える荒い海綿状の構造や、椎体の側面にぽっかりと開いた含気腔は鳥盤類にはまったく見られないものだったのである。
ハッチャーはこの椎骨とよく似たものを現在の職場――カーネギー博物館で見たことがあった。マーシュの助手時代からの付き合いで、カーネギー博物館で先に働いていた義兄のピーターソン(ハッチャーのカーネギー博物館への移籍にはピーターソンの口添えもあったようだ)が、1902年にワイオミングはランス層で発見した巨大な獣脚類の部分骨格CM 1400である。この巨大な「ドリプトサウルス属のなにか」の椎骨は、マノスポンディルスと酷似していたのだった。
ハッチャーがCM 1400の研究をどこまで本気で進めるつもりがあったのかは、今となっては謎である。とはいえトリケラトプスやハドロサウルス類(つまるところエドモントサウルス)の天敵としての巨大なドリプトサウルスというところにはこだわりがあったようで、1904年には来る角竜のモノグラフ用にトリケラトプスとハドロサウルス類、ドリプトサウルスを描いた生態復元画をナイトに依頼した。ドリプトサウルスについては骨格の不明な部分が相当にある(この時点ではまだランベによるカナダ産頭骨の再記載は出版されていなかった)ことから、ハッチャーは植生でうまくカバーするようナイトに注文している。しかしながらハッチャーは結局ドリプトサウルスをあきらめ、かわりにもう1匹のトリケラトプスを描いてもらったのだった。
ナイトの復元画の納入を見届けたハッチャーはほどなく体調を崩し――遺伝性の疾患に苦しんでいたハッチャーにとって、それ自体は珍しいことではなかった――、書きかけの角竜のモノグラフと首なしのディプロドクスを残して帰らぬ人となった。角竜のモノグラフ、そしてランス層産巨大ドリプトサウルスの一報とマノスポンディルスの再記載は1907年になるまで出版されることはなかったのである。

↑“ディナモサウルス・インペリオススDynamosaurus imperiosus”のホロタイプNHMUK R7994(旧AMNH 5866;上段左)、CM 1400(上段中)、“マノスポンディルス・ギガスManospondylus gigas”のホロタイプAMNH 3982(上段右)、ティラノサウルス・レックスのホロタイプCM 9380(旧AMNH 973;下段)。スケールバーは1m。
NHMUK R7994とCM 1400は首から後ろの要素もある程度発見されている。
20世紀になり、化石収集競争の先頭に立ったのはオズボーン――打倒マーシュを掲げた点においてコープの精神的後継者――率いるAMNHの古脊椎動物学部門であった。そしてそこにさらなる新興勢力――鉄鋼王カーネギーの潤沢な資金を背景にそびえるカーネギー自然史博物館と、シカゴの豪商マーシャル・フィールドが設立したシカゴ・コロンビアン博物館(現フィールド博物館:FMNH)が追随することとなったのである。研究だけでなく「展示」(そしてそこから続く教育)への強い関心があったオズボーンにより、「化石戦争」は今日まで続く新しい形のものへと変貌したのであった。
AMNHの展示を拡充するうえでオズボーンが頼りにしていた化石ハンター(ただしマーシュの助手だったころのハッチャーのような不安定な立場ではなかった)が、言わずと知れたバーナム・ブラウンである。プリンストン大学時代のハッチャーそしてピーターソン(ハッチャーにプリンストン大へ引き抜かれる前にはAMNHで化石ハンターをしていた;後釜がブラウンである)と共に「化石人間」として1898年からパタゴニア遠征調査を行ったブラウンは、「一人置き去りにされた」(引き続き化石調査を自由に続けたかったブラウンが合意のもと隊を離脱したというのが実際のようだ)のち、1900年に帰国するとすぐに西部へと派遣された。喫緊の目標は、展示映えする白亜紀の恐竜の頭骨――この当時、事実上YPMとUSNM、そしてカンザス大(学生時代のブラウンが発掘に携わったもの)にしかなかったトリケラトプスの頭骨の確保である。
(ブラウンは学生時代にフィールドで、プリンストン大時代のハッチャーと一度遭遇している。パタゴニアの遠征中、ブラウンはハッチャーのときおり棘のある性格に触れつつも、かなりの好印象を持っていたようだ。ピーターソンと3人で遭難しかけ、一つしかないベッドで二日間寒さをしのいだというエピソードもある。)
コープの一連のモリソン層産コレクションを受け入れたうえ、ボーン・キャビンでの発掘を精力的に行っていたAMNHは、ジュラ紀のコレクションで他の博物館に対して優位に立っていた(カーネギー発掘地の発見によってひっくり返されるのだが)。したがって、「マーシュ越え」を狙うオズボーンとしてはなんとしてでも白亜紀の(それもできるだけ展示映えする)恐竜化石が欲しかったのである。
かくしてワイオミングはランス層を訪れたブラウンは、ハッチャー隊によって散々ほじくり返されたエリアを避けて発掘を行った――が、“クラオサウルス”(現エドモントサウルス・アネクテンス;AMNH 5863)の部分骨格こそ見つかれど、トリケラトプスの化石は角の破片がせいぜいであった。しかしブラウンはここで伝説の第一歩をしっかりと踏み出していた。1900年10月、ばらけた大型獣脚類――マーシュの記載に一致するものはないようだった――の骨格に出くわしたのである。
この骨格の関節の大半は外れていたが、頸椎は部分的に関節した状態にあった。周囲には大小さまざまな皮骨が散らばっており、ブラウンは一見してケラトサウルスの類縁と見て取った。骨格はハドロサウルス類や“パラエオスキンクスの歯”(すでにイグアナではなく鎧竜を指す言葉となっていた)、トリケラトプスのフリル、魚鱗やその他の骨片と混在した状態にあり、ブラウンにはそれらがこの肉食恐竜の最後の食事のあとかたであるように思われた。
(実のところ、ティラノサウルスとの出会いはブラウンにとってこれが初めてではなかったようだ。カンザス大時代に発掘に携わった標本の中には、KU 1357として知られるティラノサウルスの単離した趾骨がある。)
かくしてブラウンは未知の大型獣脚類――AMNH 5866をニューヨークへと送り出したが、結局1900年のこの調査では保存のよいトリケラトプスの頭骨を持ち帰ることはできなかった。翌1901年はスミソニアンの三畳系の調査に「貸出」されることとなったブラウンだったが、ツキは1902年に巡ってきた。
ブラウンは様々な時代の様々な動物化石の発掘経験をすでに相当に積んでいたが、結局彼の心をもっともつかんだのは恐竜であるようだった。ランス層でのトリケラトプス探しの機会を再び与えるようオズボーンに催促していたブラウンだったが、一方でさすがのオズボーンもAMNHの展示の拡充ばかりにかまけているわけにもいかず、自らの研究のために新生代の哺乳類化石をもっと採集する必要を感じていた。
ブラウンのリベンジの機会は思いがけず訪れた。ブロンクス動物園の園長兼学芸員であったホーナデイ(アメリカの自然保護活動を牽引する一方、オズボーン以上に人種差別で悪名高い)が、オグロジカの生態調査中にモンタナ州はヘルクリーク周辺でトリケラトプスの化石のうわさを聞き付けたというのである。
ホーナデイはブロンクス動物園の運営団体の理事長であったオズボーンと親交があり、同行していた写真家のハフマンに撮らせた化石の写真をオズボーンとブラウンに見せた。果たして、そこに写る化石のいくつかはトリケラトプスのものに違いなかった。
コープとマーシュの「化石戦争」において、白亜系の主戦場はモンタナのジュディス・リバー層、ワイオミングのランス層にコロラドのデンヴァー層であった。モンタナやサウスダコタ、ノースダコタにはジュディス・リバー層よりも新しい、ランス層やデンヴァー層の同時異相――しばしば「大褐炭層」と呼ばれてきた地層があったが、そこではコープ隊がわずかな化石を採集していた程度だった。
つまりこの地層(実のところ「大褐炭層」の下位層)――ヘル・クリークHell Creek層(ブラウンの命名による;マーストリヒチアン後期;約6720万~6604万年前)は、「化石戦争」の戦禍をほとんど被っていない地層であった。かくしてブラウンはリチャード・スワン・ラル――オズボーンの学生だった――を相棒に、バッドランドへと旅立っていったのである。
地元住民は一帯に化石がたくさん転がっていることをとっくに知っており、ブラウンは運が向いてきたのを感じていた。ブラウンとラルはホーナデイの旧友であるシーバーがかつて所有していた牧場にたどり着き、その周辺でついに立派な“ステルロロフス”の頭骨(と四肢の一部;AMNH 970)を発見したのである。
そうこうしているうちにオズボーンからの𠮟責の手紙(以前ブラウンが採集した標本について、プレパレーションし損だったという内容である。母岩の付いたままの大きな化石の評価をフィールドで下すのは今も昔も難しい)が届き、ブラウンは謙虚に反省した――が、名誉挽回のチャンスは目の前にあった。ヘルクリークを挟んでシーバーの避難小屋の向かいにある崖で、ブラウンは再び、「マーシュの記載にはない」巨大な獣脚類の化石を発見したのである。
化石は急峻な崖の途中に埋まっており、馬が入るのもおぼつかない場所であった。しかも化石は青い砂岩のコンクリーション――ブラウンがそれまで経験したことのないほど硬い岩に覆われていたのである。最寄りの鉄道駅までは200km以上あり、掘り出した化石をどうやって駅まで運ぶかも問題であった。ダイナマイトを駆使しての発掘が、始まった。
困難な発掘の中で夏から秋へと季節は移ろい、降雪の時期が迫りつつあった。ブラウンに相棒としてつけられたラル――のちにアマースト大学を経てイェール大学へ移り、ハッチャーの角竜のモノグラフを完成させ、教授としてキャリアの最後まで在任した――は年齢も学歴もブラウンより上だったが、もちろんフィールドでの経験はブラウンの方が豊富であり、したがって調査の隊長はブラウンであった。こうした背景からふたりの関係は微妙な緊張をはらんでいたが、うまく調査を回していた(ブラウン自身、調査中にラルの評価をよい方向に改めたことを記している)。しかし長引く発掘を前にオズボーンから預かった路銀は底を突こうととしており、吹雪の季節は厳然として立ちはだかっていた。ラルをまずニューヨークへ帰し、それから200km近く離れたマイルズシティへ化石を次々と運び出した。駅にはオズボーンのよこした空の有蓋貨車があり、概算で4.5~6.8tほどの(母岩付きの)化石はかくしてニューヨークへと送り届けられたのであった。
ブラウンが発見した巨大な獣脚類の化石――AMNH 973は1900年に発掘されたAMNH 5866とはまた違ったやっかいさを(はるかに上回るレベルで)抱えていた。AMNH 5866は母岩も化石も随分もろかったのだが、AMNH 973は(発掘中から明らかだった通り)ひたすらに硬いコンクリーションがクリーニングを妨げていたのである。
一足先にニューヨークへ戻っていたラルと、AMNHが当時世界に誇るプレパレーターであったピーター・カイゼンとポール・ミラーがAMNH 973のクリーニング(母岩ごと化石を粉砕してから再接合するテクニックも駆使されたようだ)を進めていく中で、もう一つの深刻な問題が頭をもたげてきた。AMNH 973の部分的な骨格の保存状態は抜群によかったのだが、しかしその割に妙に部分的にすぎるように思われてきたのである。
1905年のフィールドシーズンが近づくころ、オズボーンはAMNH 973――デイノドンの新種(オズボーンはコープの教え子ではあったが、分類についてはもっと真っ当な見方をしていた)とみられる標本と、それからAMNH 5866の記載の執筆にとりかかり始めていた。論文の執筆を進めるうち、AMNH 973の発掘場所の再訪は避けられないというところでオズボーンとブラウンの意見は一致した。掘り残しがある可能性をふたりとも感じていたのである。かくして1905年5月、ブラウンは(前年に結婚した妻のマリオンをこっそり料理係として連れて)再び現地へ舞い戻ることとなった。
案の定、AMNH 973の掘り残しはそこでブラウンを待っていた。
AMNH 973を掘りきるためには崖を相当に(ダイナマイトで)掘削することが必要であり、追加要素のクリーニングも含めれば年単位で時間を要する気配があった。腰を据えてかかるのが一番だったが、オズボーンたちには気がかりなことがあった――見えないタイムリミットが近づいていたのである。
AMNH 973の産地を再訪した際、情報収集に出たブラウンはカーネギー博物館のウィリアム・アッタバック(ピーターソンと同様にイェール時代のハッチャーの助手を務めており、カーネギー博物館でも3人なかよく働いていた)のチームが数週間前に一帯を調査していたことを知った。幸い現場は3年前のままだったのだが、CM 1400――ハッチャーの死から1年ほどしか経っておらず、角竜のモノグラフは未完のままだったため、この標本に関する情報は寝耳に水だった――のプレパレーションが最終段階にあり、まもなく記載されるであろうという一報までニューヨークに届いていたのである。
CM 1400には頭骨まで含まれているという話で、であればホランド麾下の(キャリアのどこかでAMNHでの経験があった)研究者たちはそつなく新種として記載するであろうことは容易に想像がついた。そしてAMNH 5866(この時点では歯骨だけが図版の制作のために優先的にクリーニングされていただけだったらしい)には頭蓋はさっぱり残っておらず、AMNH 973の頭骨は破片しか出てこなかったのである。CM 1400の実態はいまだ不明であったが、現状でAMNH 5866とAMNH 973が「勝てる」見込みはあまりなかった。CM 1400――「ドリプトサウルスの新種」とAMNH 973・5866――「デイノドンの新種」は同じ分類群の可能性が多分にあり、であれば先に命名された方が先取権の問題で圧倒的に有利だったのである。
オズボーンはAMNH 973とAMNH 5866の記載論文をデイノドン類――白亜紀の北米の大型獣脚類の集大成として仕上げようとしていた。焦る仕事ではなかったはずなのだが、カーネギー博物館に先を越されてしまえばそれまでの苦労が水の泡になることは目に見えていたのである。「マーシュの記載していない」巨大獣脚類の名付け親がカーネギー博物館になることは認めがたいことであった。
悪天候で現地への到着が遅れたこともあり、必死の思いで掘り進めたブラウンと助手のレロイ・パーキンは、ほどなく頭骨と思しき巨大な化石を含んだコンクリーションにぶちあたった。しかしこれは腸骨で、本物の頭骨の残りをすぐ採集して持ち帰るというわけにはとてもいきそうになかった。クリーニングも考えると、時間はもはや残されていないように思われたのである。
(ブラウンはフィールドでの印象からAMNH 973とAMNH 5866が別物であるとかたくなに信じていた。AMNH 973の掘り残しの発掘の最中にも別属として記載するようブラウンから手紙で促されているあたり、オズボーンは必ずしもそうではなかったようである。ティラノサウルスの原記載のためにウィリアム・ディーラー・マシュー(もともとオズボーンの助手で、ブラウンと親しかった。画才もあり、ナイトの復元画の監修役も務めていた。エドウィン・コルバートの義父でもある)の描いた骨格図の原画(つまりAMNH 973を描いたものである)には「デイノドン」の文字とともに「背中と脇腹は不規則な小さい、インチ厚の骨板で覆われていた」との文が添えられている。オズボーンはマシューに骨格図の制作を依頼した時点では、AMNH 973とAMNH 5866を同じ分類群――デイノドンの新種として記載するつもりであったようだ。フィールドでの観察に基づくブラウンの意見――両者をかたくなに別属と主張した――を容れて、最終的な論文としたように思われる。)
ブラウンがAMNH 973の発掘を今度こそ終えて数か月後、クリーニングの完了を待たずしてオズボーンはAMNHの紀要にてティラノサウルス・レックスTyrannosaurus rex――AMNH 973をホロタイプとする新属新種の獣脚類を記載した。オズボーンは土壇場で(7月下旬までに)、この恐竜を新属とする決意を固めたのである。結局頭骨のクリーニングは間に合わなかった(論文中に明記までしている)が、相当量が保存されており、いざとなればCM 1400に対して先取権を主張できると踏んでのことであったようだ。オズボーンはティラノサウルス・レックスに続いて、AMNH 5866をホロタイプとしたディナモサウルス・インペリオススDynamosaurus imperiosusを命名した。この時点でティラノサウルスとディナモサウルスを分かつ特徴は、皮骨の有無だけであった。
オズボーンはまた、それまでに命名された北米産の白亜紀の中~大型獣脚類についても分類の整理を行った。コープによって事実上デイノドンと合体させられていたドリプトサウルスを(アメリカ東部というはるか遠くで発見されていたことから)ドリプトサウルス・アクイルングイスに限定し、それからライディによる原記載を尊重する形で(つまり異歯性を重視して)デイノドンを復活させてアウブリソドンをジュニアシノニムとした。そして、エドモントン層(現ホースシュー・キャニオン層)がジュディス・リバー層よりも新しい時代であり、かつニュージャージーとも遠いことから、ランベがドリプトサウルス・インクラッサトゥスとして記載したCMN 5600をホロタイプ、CMN 5601をパラタイプとして新属新種――アルバートサウルス・サルコファグスAlbertosaurus sarcophagusを命名したのである。オズボーンの観察によれば、アルバートサウルスの歯骨歯の本数や厚さはデイノドンとディナモサウルスの中間にあるようだった。
1905年12月3日、NYタイムズ日曜版にAMNHの恐竜発掘を紹介する全面記事が掲載された。ブラウンにとって初めての新聞沙汰であり、見出しの真下――センターには、マシューの描いたティラノサウルスの骨格図が大きく掲げられていたのである。化石の展示はまだだったが、かくして「百獣の王」のキャッチフレーズをひっさげ、ティラノサウルスは大衆社会へのデビューを果たしたのであった。
(オズボーンによるティラノサウルスの原記載は「ティラノサウルスと他の白亜紀の肉食恐竜」と題されたが、この論文の出版された日は厳密にははっきりしない。英語圏でも1905年10月5日とされることが多いようだが、紀要に記された別刷りの発行日は10月4日となっている。そもそも、記された発行日と実際に印刷物にアクセスできるようになった日が一致するとは(今日でも)限らず、この時代の論文では往々にして派手にずれることもある(コープに至ってはそのあたりを明らかに不正利用していた気配がある)。日本時間の10月5日に祝っておくというのは悪くない落としどころであるようだ。コープやマーシュと比べると、クリーニングが追い付いていなくてもよいのでとにかく原記載に一般受けするビジュアルを添える、というオズボーンの現代的な発想の光る論文でもある。マシューによって描かれた原記載の骨格図は、実のところ本文とあからさまに矛盾するレベルで巨大に描かれていた。)

↑CM 9380(旧AMNH 973)の骨格図。スケールバーは1m。
今日の復元(上)と1906年ごろの復元(下)とで仙前椎の様子が変わらないである点に注意。
冬――フィールドへ出られないシーズンの間にAMNH 973(とAMNH 5866?)のクリーニングは着実に進み、きちんとした論文用の図版(原記載時のディナモサウルスの図版はアーウィン・クリストマンが担当し、続いての再記載の図はリンジー・モリス・スターリングが担当した。一方で、クリストマンはその後のモノグラフを想定してか陰影付きの未出版の図を相当数製作してもいた)の制作までこぎつけた。カーネギー博物館に対して先手を打つことに成功した(とうとう今日に至るまでCM 1400のきちんとした記載は出版されなかった)ことで、本来意図していた大著にふさわしい準備に落ち着いて時間を割けるようになったのである。
オズボーンの焦りは、結果的にいささか裏目に出てもいた。クリーニングが終わってみれば、AMNH 973とAMNH 5866を別属はおろか別種としてみることもできそうになかった――産出部位の重複する部分はことごとくそっくりだったのである。両者を分かつものは、結局のところ皮骨――オズボーンはこれがAMNH 5866のものなのか疑問を抱くようになっていた――だけだった。
かくして1906年の7月30日付で出版された再記載で、ティラノサウルスの実態――依然として未発見の部位は相当にあったが、とはいえAMNH 973とAMNH 5866を組み合わせることでかなりの部分が埋まった――が明らかにされた。ディナモサウルス・インペリオススはかくしてティラノサウルス・レックスのジュニアシノニムとなり(ホロタイプの発見は前者の方が早かったわけだが、原記載における扱いは後者の方が早かった;論文のタイトルからして後者が「本命」であったことを示している)、ティラノサウルス・レックスの標本リストにはさらにもう1標本――AMNH 5881(現NHMUK R7995;いくつかの腹肋骨と左右の後肢の大部分)が追加されたのである。
クリーニングが終わってみればAMNH 973の頭骨はそれなりの部分が非常によく保存されており、骨盤はほぼ完全に、後肢も大部分が残されていた。肩甲骨も残っており、上腕骨――AMNH 973に属するのか否かでオズボーンとブラウンはかなり揉めた(オズボーンをして原記載の出版を遅らせるほどだった)が、結局のところこの中空――つまり獣脚類には違いない――の骨は、肩甲骨の肩関節窩に非常にうまくはまったのであった。そしてAMNH 5866は頸椎がほぼ完全に残っており、AMNH 973の胴椎と合わせると仙椎までの様子をかなりはっきりと描き出すことができた。完全な骨格が知られているとはとても言えなかったが、しかしティラノサウルス・レックスは白亜紀の大型獣脚類としてはもっとも完全な骨格が知られていることも確かだったのである。
スターリングは1/40スケールで、前年にマシューが描いたごくごく概略的な骨格図よりもはるかに精緻に描き込まれたものを描き上げた。そしてオズボーンは、コープが1866年に設立したデイノドン科に特に触れることなく、ティラノサウルス科を設けたのである。
オズボーンを最後まで悩ませたのはAMNH 5866の皮骨であった。なにしろAMNH 973では奇妙なことにひとかけらの皮骨さえも発見されなかったのである。AMNH 5866はトリケラトプスやハドロサウルス類の破片と共産したのであるが、とはいえ(トリケラトプスの体を覆う皮骨についてはマーシュが報告したりもしていたのだが)こうしたタイプの皮骨はそれらの恐竜では見られないものであった。皮骨が明らかな獣脚類の肋骨と近接して発見されたこともあり、オズボーンは(最後まで懐疑的ながらも)ブラウンの意見――AMNH 5866は皮骨で装甲されている――を受け入れたのである。オズボーンは、こうした皮骨の役割は種内戦闘からの保護以外には考えにくい、と述べてこの論文の締めの文とした。
(「ティラノサウルスの皮骨装甲」についてきちんとした記載や配列の検討がなされることはとうとうなかった。ブラウンは1908年にアンキロサウルスを記載した際に「ティラノサウルスの皮骨」について触れ、アンキロサウルス(のホロタイプ)のそれと比べて表面がより滑らかで、やや不規則な形態であること、おそらくはより皮膚の深い部分に位置していたであろうことを述べている。また、「トリケラトプスの皮骨装甲」は結局のところトリケラトプスの骨格と共産したものではなく、フィールド調査でも一度も確認できなかったことにも触れている。「ティラノサウルスの皮骨装甲」が真剣に検討されることはそれからなかったのだが、2004年にカーペンターによって初めてきちんと記載・図示され、単にアンキロサウルスの皮骨であることが示されて今日に至っている。「トリケラトプスの皮骨装甲」は少なくとも角竜のものではなく、よく目立つスパイクのクラスターはパキケファロサウルス(ないしスティギモロク)の幼体のものであった。)
1906年12月30日、NYタイムズにティラノサウルス・レックスのAMNHでの展示開始を知らせる記事が掲載された。まず展示されたのはAMNH 973の巨大な下半身――ほぼ完全な状態で保存されていた仙椎と骨盤、そして相当部分が保存されていた左右の後肢にアーティファクトを加えて組み上げたものであった。優雅な足取りでマウントされた脚の間には現生鳥類の骨格がちょこんと立ち、来館者に両者の類似を訴えかける仕掛けになっていたのである。記載を前倒ししてまで急いで命名したにもかかわらず、そこからわずか1年で(部分的とはいえ)オズボーンは写真映えするマウントの展示にまでこぎつけたのであった。ナイトによる復元画がこれに花を添え、AMNHは、名実ともに「マーシュの記載していない」白亜紀の巨大獣脚類を巡る競争に勝利したのである。
(この時点では、ティラノサウルス・レックスとして同定された第III中足骨は存在しなかった。オズボーンはティラノサウルスの第II、第IV中足骨が(ジュラ紀の大型獣脚類とは異なり)第III中足骨と深く嚙み合う構造であることを見抜いてはいたが、“オルニトミムス・グランディス”との類似には気付いておらず、従ってAMNH 973の足の甲は実際よりもやや幅広のものとしてマウントされている(末節骨も実際よりは大きめのものが据えられた)。このマウントは後述のAMNH 5027のマウントの完成後もしばらく常設展示に留まっており、 しばらくは同じフレームに入れて撮影することが可能であった。開始のタイミングははっきりしないものの、大量のアーティファクトと共にマウントされたAMNH 973の復元頭骨も展示されており、やはりAMNH 5027のマウント・実物頭骨と共に展示されていた時期もあったようである(写真左端、ティラノサウルスの脳函(AMNH 5029とAMNH 5117)と共に写っている)。この復元頭骨はスターリングの図とは側頭窓の作りが別物で、再記載の図版とは制作のタイミングがずれていたようにも思われる。これらAMNH 973の初期の展示物は当然現存していないが、カーネギー博物館のリニューアルまではCM 9380の復元骨格の頭部(と足元の実物頭骨)は「旧復元」のままであったし、骨盤や後肢の細かな破損・欠損部を充填したアーティファクトは今日までAMNH製のものが残っている。)
1908年夏、ブラウンは1年ぶりのフィールド――ふたたびモンタナのヘル・クリーク層へと赴いた。今度の相棒は腕利きプレパレーターのカイゼンである。1906年の予備調査で発見されたハドロサウルス類の採集をひとまずの目標に、前回のヘル・クリーク沿いから50kmほど東、ビッグ・ドライ・クリーク沿いで調査が行われることになっていた。実のところ、オズボーンの求めるものはティラノサウルスの追加標本であった。
現地で牧場を営むトウィッチェル一家と仲良くなりつつ、いざ7月1日に現地に到着すると、ブラウンは予備調査で発見されていたハドロサウルス類の採集をすっぱりあきらめた。――が、ここでブラウンは生涯最高の化石との出会いを果たしていた。キャンプへの道すがら、風化した4個の尾椎が転がっているのを見つけたのである。
なんということはない尾椎のようだったが、ちょっと掘ってみると――化石はAMNH 973の時のようなコンクリーションではなく、固結度の低い砂岩に埋まっていた――状況は一変した。そこにあったのは、小さな丘へと続く15個の尾椎だったのである。そして、尾椎はこれまでにブラウンが見たことのない形態であった。
尾椎を追いかけてキャンプを丘のそばへ移したブラウンは、7月3日にティラノサウルスと思しき頭蓋と下顎――尾椎の主が埋まっているのを見つけた。お祭り男のブラウンにとって、独立記念日前日のこの出来事は吉報もいいところであった。近隣(日本の感覚にはあてはまらない)住民とともにトウィッチェル牧場で独立記念日を派手に祝った一行は、尾椎の残りが丘の中――砂岩のコンクリーションへと続いていることを確認したのである。
料理人がろくに仕事をしなかったことを除けば、発掘は極めて順調に進んだ。コンクリーションはAMNH 973の時ほど硬いわけではなく、崖を上ることもないのもあって足場はずっとよかったのである。ダイナマイトで丘の上部を爆破し、7月半ばには化石の全容が明らかになった。
そこにあったのは、デス・ポーズで横たわるティラノサウルスの巨大な骨格であった。頸椎から尾の中ほどまでが完全に関節していたそれのかたわらには、完全な頭蓋と下顎まで残されていたのである。
チームはまたしてもトウィッチェル牧場で(大量のチキンとアイスクリームで)発見を祝い、オズボーンもこれをいたく喜んだ。8月の下旬に現地を訪れたオズボーンは、「頭骨だけでもこの夏の作業の価値がある」とブラウンが手紙で書いた通りのものを目にできたのだった。
発掘そのものは9月上旬までにカタがつき、10月第一週までひたすらに梱包作業が続いた。一連の調査は大成功に終わり、この骨格――AMNH 5027のほかにティラノサウルスの単離した脳函AMNH 5029やトリケラトプスの頭骨AMNH 5028、オルニトミムス類まで採集できたのである。

↑AMNH 5027(上、中)とCM 9380(下)の骨格図。スケールバーは1m。
AMNH 5027の復元骨格の最初のバージョンに基づくもの(上)では前肢が3本指であり、肩帯と四肢にCM 9380(灰白色)をそのまま組み入れられているため、今日の復元(中)と比べて四肢がマッシブかつわずかに長い点に注意。AMNH 5027では頸椎と胴椎が病変のために短くなっているが、本来CM 9380とほぼ同じサイズの個体である。
AMNH 5027のクリーニングは着実に進められ、1912年になるとオズボーンは頭骨の詳細な記載を行った。AMNH 5027の頭骨は当時知られていた獣脚類のものとしては最大かつ最高の保存状態を誇っており(完全な頭骨が知られていた獣脚類は他にケラトサウルスとアロサウルスくらいしか知られておらず、前者は変形がひどく後者は下顎がいまひとつよくわかっていなかった)、AMNHの所蔵していた一連のアロサウルスの標本と合わせて大型獣脚類の頭骨の詳細がここで明らかになったのである。マーシュの死後10年以上を経て、ついに角竜キラーたる「攻撃用に特殊化したそれらの特徴」が明らかになったのであった。
AMNH 5027の記載と並行して復元骨格の展示計画・制作も進められた。オズボーンの手元にあったティラノサウルスの標本のうちAMNH 973とAMNH 5027は驚くべきことにほぼ同サイズの動物であり、しかもAMNH 973とAMNH 5027をそれぞれのレプリカで相互補完してやれば尾の後半部以外ほぼ全身がそろってしまう格好であった。両者とも保存状態は抜群によく、最大最強最後の肉食恐竜の復元骨格をいきなり2体展示できるチャンスにオズボーンが飛びつかないはずはなかった。
(実際のところAMNH 973=CM 9380はAMNH 5027よりもわずかに大きいが、このサイズの違いは個体変異で吸収できてしまう程度ではある。体サイズはさておきCM 9380はAMNH 5027よりもずっとがっしりした、より成長の進んだ個体であり、問題はむしろこちらにある。結果的に、今日でもAMNH 5027の復元骨格は実際よりもはるかに頑強かつわずかに長い(非アークトメタターサルの)後肢で立っている。)
2体の巨大な獣脚類の骨格を展示するうえで立ちはだかる問題はいくらでもあった――単なる技術的な困難もあったが、限られた館内空間にティラノサウルス2体を展示・設置するのは極めて困難なように思われた。博物館の増築にかかるコストは言うまでもなかったが、それでも(あるいはそれゆえに)オズボーンはあきらめず、 「ティラノサウルスポージングコンテスト」を行ってAMNH 973とAMNH 5027の展示計画を練り続けた。採用されたのはブロンクス動物園の爬虫類担当学芸員レイモンド・L・ディトマーズによるもので、倒したばかりの“トラコドン”を巡って2頭のティラノサウルスが争うという構図であった。そして彫刻家としての才能にも恵まれていたクリストマンによって1/6スケールのティラノサウルスのフル可動の骨格模型が木彫りで2体(+“トラコドン”の代役としてもう1体)制作され、復元骨格のポーズ・レイアウトの検討に供されたのである(この模型は最終的にポーズを変更して常設展示に回された)。
完全度が高く、それゆえ復元骨格に組み込まれる化石の量が多くなるAMNH 5027は“トラコドン”の上に覆いかぶさった低い姿勢で、これであれば重い頭骨もそのままマウント可能であると見積もられた。完全度が低く(それゆえ、化石本体よりも大胆にアーティファクト部分に支持材を通すことができる)、しかもすでに制作された復元頭骨(石膏に実物標本を埋め込んだため、作り直しは困難だった)を流用することが前提であったAMNH 973は雲突くばかりの高さに立ち上がり、AMNH 5027から獲物を奪おうとする(ついでに、あからさまに不正確なものとなった復元頭骨は来館者から見えにくくなる)ポーズで組み立てられることとなったのである。実のところこれに近い(完全な頭骨を含むより完全度の高い骨格を低い姿勢で、そうでない方を高い姿勢でマウントする)レイアウトの展示はすでに“トラコドン”のペアで実現できており、技術的な問題はそれほど致命的ではなかったようでもある。
スポンサーへのアピールを兼ねてか、この「展示用復元骨格を制作するための新手法」は1913年にAMNHの紀要にて出版された(没ポーズの写真も残っている)が、しかしオズボーンの剛腕をもってしても、この展示計画の前に立ちはだかる経済的な障壁を乗り越えることはできなかった。かくしてティラノサウルスの展示計画は縮小され、AMNH 973は(引き続き)骨盤と後肢のマウントと復元頭骨が展示されることとなった。AMNH 5027はよりおだやかなポージングとされ、結果的に(高い位置でマウントしなくてはならなくなったため)頭骨は軽量なレプリカを用いてマウントされることとなった。
(AMNH 5027のプレパレーション中の記録(というか宣伝)写真は広く世界に紹介されたようである。日本でも大正7年(1918年)にAMNH 5027の組み立て中の写真が出版されているが、これはどうやら日本で初めて示されたティラノサウルスのビジュアルでもあるようだ。存在そのものはラルの論文の抄訳という形で大正元年(1912年)には紹介されているが、この時のカナ表記は「チラノサウルス」であった。「ティラノサウルス」表記は大正10年(1921年)にはすでに生まれており(ついでにTyranosaurusと誤記された)、わりあいに起源が古い。)
ハーマンの号令の下、チャールズ・ラングはひたすらにアーティファクト――AMNH 973の石膏レプリカをもってしても埋まらなかった部位を補完するもの――をこね続けた。危なっかしい高所作業の連続の末、1915年の10月についにAMNH 5027の復元骨格は完成した。人類時代展示室――化石ホールにすぐに置ける場所などなかった――の天井すれすれにそびえ立ったそれは、ティラノサウルス・レックスが真に恐竜の王としてよみがえったことをはっきりと示していたのである。オズボーンは1916年にそれまでの獣脚類に関する研究の集大成を出版し、AMNH 5027のありようをクリストマンに存分に描かせ、アンダーソンによる復元骨格の写真で論文を飾らせたのだった。
(オズボーンはこのとき初めてAMNH 3982――マノスポンディルス・ギガスについて言及し、ハッチャーの意見――マノスポンディルスが角竜などではなく獣脚類、それもおそらく「巨大ドリプトサウルス」すなわちティラノサウルス・レックスのものとする意見を追認した。オズボーンはこれがティラノサウルスの第10頸椎ないし第1胴椎に酷似していることを見抜いたが、保存のよくない(ゆえに粗い海綿状の組織がむきだしになっていた)単離した椎骨に過ぎないそれに基づく学名を疑問名とみなし、捨て置くこととした。ラーソンはBHIが2000年にサウスダコタで発掘を始めた(前年の終わりにバッキー・ダーフリンガー(言うまでもなく“バッキー”の名は彼にちなんでいる)が一部の骨を持ち込んだ)ティラノサウルス・レックスの標本BHI 6248(E.D.コープというそのまんまな愛称がついている;相当な大型個体であり、AMNH 3982と同程度のサイズには違いない)がAMNH 3982の掘り残しである可能性を示唆した。BHI 6248は部分骨格に過ぎないが、とはいえ上下の顎をはじめとする要所が残っており、これがAMNH 3982の掘り残しであるならば、マノスポンディルス・ギガスは分類上有意な特徴を残した標本をホロタイプとしているという格好になり、ティラノサウルス・レックスをジュニアシノニムにできうる(ので、BHI 6248は学術的に意義深い)――というのがラーソンの言い分であった。BHI 6248がAMNH 3982の掘り残しである、とする根拠は実のところダーフリンガーによる発見以前のいつごろかに誰かが椎骨を掘り起こして脇に積み上げていたらしい、という以上の話ではなく(ラーソンは産地の大まかな位置(「サウスダコタ州モロー川の南側」)も根拠に挙げているが、そもそも先述の通りコープはAMNH 3982の産地情報を原記載に一言も残しておらず、ハッチャーにしても口伝で「サウスダコタ産」とまでしか聞き出せていない。ラベルにこのあたりの情報がなかったのは言うまでもない)、もし本当にBHI 6248がAMNH 3982の掘り残しであったとして、そもそもマノスポンディルス・ギガスは動物命名規約第4版(2000年~)いうところの遺失名nomen oblitumの条件を満たしている(ので、強権発動もなにもないまま、ひとたび正式な形で遺失名であると述べてしまえば終わりとなる)格好である。結局のところBHI 6248をAMNH 3982の掘り残しとみる意見は真剣に検討されたことがなく、マノスポンディルス・ギガスは遺失名としてきちんとした出版物で宣言されることもなく単なる疑問名のまま今日まで至っている。)
それから30年近くの間、AMNH 5027はティラノサウルス・レックス唯一の全身復元骨格として、化石ホールにようやく移設された1917年以降はホロタイプ――AMNH 973を従えてずっと一人で立ち続けた。ほどなくブラウンは妻を病で亡くし、ブラウンらAMNHの調査隊とGSCに雇われたスターンバーグ一家(長男のジョージは当初AMNH隊に雇われていた)による第二次化石戦争によって姿を現したはるかに完全な「デイノドン科」――ゴルゴサウルス・リブラトゥスの発見にともなってAMNH 5027は1927年に手の第III指をもぎ取られた。
1908年からずっと理事会会長としてAMNHに君臨していたオズボーンは1934年に退職し、名誉職にありつつ静かに余生を過ごすはずだった――が、翌1935年に書きかけのモノグラフを残して机に向かったまま心臓発作で息を引き取った。館長としてオズボーンの精神的後継者となったアンドリュース――実態がどうあれ、ブラウンとはライバル関係にあった――にも世界恐慌から来る博物館の財政難はどうすることもできず、AMNH――ひいては恐竜研究はゆっくりと時代の波に取り残されていった。シンクレア石油からのフィールド調査への資金援助を取り付け、ジョージ・ゲイロード・シンプソンやエドウィン・ハリス・コルバートといった新星を迎えていたAMNHであったが、資金難は研究能力を着々と奪いつつあったのである。
1941年、古脊椎動物学部門創設当時からのメンバーであり、ブラウンとともに共同で部門の代表を務めていたウォルター・グレンジャー――正式な学位を長年持たないまま、17歳から半世紀以上にわたってAMNHにあり続けた生粋の叩き上げ――は野外調査中に倒れ、帰らぬ人となった。長年の仕事仲間を失い、代表として一人残されたブラウンだったが、それでも前に進み続けるしかなかった――第二次世界大戦の影が色濃く落ちる中で、疎開を口実に古脊椎動物学部門の予算状況を少しでも改善しようとしていたのである。オズボーン亡き今、ブラウンはカーネギー自然史博物館にAMNH 973――カーネギー自然史博物館を出し抜くため、論文の出版を強行してまで命名されたティラノサウルス・レックスのホロタイプを売却しようとしていたのだった。
1940年の12月から続いたカーネギー自然史博物館との交渉はうまくまとまり、1941年の暮れにAMNH 973(とAMNH 5029から作成された脳函エンドキャスト)は15個の木箱と4つの紙箱に詰め込まれ、トラックでニューヨークからピッツバーグへと旅をした。かくしてなけなしの7000ドル(今日の価値で15万ドルあまり――化石市場の異様な状況を見ればほとんどギャグのような安さである。運賃は今日の価値で2300ドルほどかかり、カーネギー自然史博物館の理事が負担した)が古脊椎動物学部門の懐へと転がり込んだのである。
(老境のブラウンとラングがティラノサウルス・レックスのホロタイプを点検している写真はアーカイブによれば1942年8月撮影とされており、であればカーネギー自然史博物館でマウントされる直前のタイミングと思われる。AMNHで長らく展示されていた復元頭骨と骨盤・後肢のマウントはそのまま台座ごと売却されているのだが、椎骨をはじめとする残りの部分のアーティファクトはカーネギー自然史博物館で一から制作されたということのようだ。)
1942年、AMNH 973あらためCM 9380の受け入れから1年足らずでカーネギー自然史博物館に新展示――2体目となるティラノサウルス・レックスの復元骨格が加わった。カーネギー博物館がほぼ40年がかりで念願の標本を手に入れた一方で、AMNHの研究部門を取り巻く状況はさらに厳しくなっていった。予算はさらに削減され、定年制が導入され、そしてブラウンは1942年の夏にとうとうAMNHの第一線を退くことになったのである。
(ブラウンはこののち、AMNHの名誉キュレーターでありつつも、戦略情報局と戦時経済局での任務(1941年の春にアンドリュースは戦争協力に関する政府の緊急要請を館の要職にあった古生物学者に通達していた)や(それ以前にも副業としてよくこなしていたが)石油探鉱のコンサルタントとして働いた。特に地中海周辺での豊富なフィールド経験を戦略情報局と戦時経済局は高く買っていたというのだが、70代を目前にあやうくハニートラップに引っかかりかけたというエピソードもある。)
戦争が終わってもなお、AMNHからティラノサウルスは去り続けた。1960年になり、AMNH 5866――ディナモサウルス・インペリオススのホロタイプはAMNH 5881もろともBMNH(現NHMUK)――大英自然史博物館へと売却されたのである。生まれて初めて大西洋を渡ったこれらの標本はAMNH 5027のレプリカ(この場合、CM 9380のレプリカ部分も含まれている)と組み合わされ、恐竜ギャラリーにてウォールマウントとして展示されたのであった。
(かくしてAMNH 5866はBMNH R7994(現NHMUK R7994)、AMNH 5881はBMNH R7995(NHMUK R7995)となった。AMNH 5866の皮骨はどこかしらのタイミングで分離され、現在ではNHMUK R8001となっている。)
期せずしてブラウンとオズボーンらが手塩にかけてきたティラノサウルスたちの集合体となったこの復元骨格だったが、怪我の功名とでもいうべき出来事に見舞われていた。これの組み立て・展示にあたったニューマンは、最低限の労力でできるだけ最新版に近いティラノサウルスの復元骨格の制作を試みた――ゴルゴサウルスのホロタイプの発見により、とうの昔に長すぎる復元であることが明らかだったAMNH 5027の尾はいくつかの椎骨が抜き取られて短縮されたが、「新規造形」が必要となるCM 9380の足は非アークトメタターサルのまま残された――が、ここで恐竜ギャラリーの構造がネックとなった。アーチ屋根のぶん天井は高かったのだが、壁際の高さはそれほどでもなく、全身を立体的にマウントするには手狭な空間だったのである。
ニューマンはここで、AMNHのマウントの状態では後頭顆が頸椎にうまく関節しないこと、頸椎の形からして首は鳥のように背側に引いた状態が基本であるらしいこと、ゴルゴサウルスの尾の後半部(言うまでもなくティラノサウルスでは未発見だった)の前方関節突起がよく伸長しており、尾の左右方向への可動性が制限されていることを見て取った。恐竜の発達した血道弓は尾を引きずる際の保護という文脈で語られることがよくあったが、結局のところもっともよく発達した血道弓は尾の近位部――引きずりようのない部分に並んでいたのである。獣脚類の行跡に尾を引きずった痕跡がみられることはめったになく、尾の後半部の左右方向への可動性が制限されていることも鑑みると、ティラノサウルス――ひいては獣脚類が尾を地面に引きずって歩く(実のところこうした復元画が20世紀前半に描かれることはそうなかった)、あるいは地面に向かって垂らしていたとはとても思われなかったのである。走鳥類のように大腿骨を水平近くに保ち、膝下だけを主に使って移動したとも思われなかったが、しかしティラノサウルスにほどよい長さの(そしてトカゲのようにくねくねと動くわけでもない)尾があるのなら、前半身とのバランスは十分に取れるように思われた。ティラノサウルスは――同様のボディプランを持つ獣脚類は鳥のように上半身をおおよそ水平に保って歩いていたのである。
アラン・チャリグとの確執を残しつつ、かくしてほぼ水平姿勢で腹肋骨までマウントされたティラノサウルスのウォールマウントが、ロンドンはサウス・ケンジントンでおひろめされた。1970年にニューマンは論文を出版し、同じ年にラッセルはより過激なポージングで描いたダスプレトサウルスの骨格図を解き放った。恐竜ルネッサンスはすでに始まっていたのだ。
それから半世紀以上が過ぎたが、相変わらずティラノサウルス・レックスは元気に過ごしている。AMNH 5027はAMNHのリニューアルに合わせて解体され、まだ残っていた母岩を完全に取り除かれたのちに現代的な保護剤で補強・修復された。頭骨を新レプリカに交換のち尾を切り詰められた(後肢はそのまま残った)マウントは、つつましげなポーズで来館者を迎えるようになってずいぶん経っている。大英自然史博物館のウォールマウントは解体されたが、その一部――“ディナモサウルス”の顎はこの恐竜の数奇な運命を伝えている。そしてCM 9380は石膏の山の中から頭骨を「救出」されたのち、因縁のCM 1400を足元に従え、オズボーンがとうとう果たせなかった夢――もう一頭のティラノサウルス(MOR 980のレプリカ)とともに獲物を奪い合う姿で組み直されたのだった。
保存状態がよく、完全度もはるかに優れた標本がいくつも知られていることにより、CM 9380の分類学的な価値は長らく顧みられることがなかった。しかし昨今のティラノサウルスを巡る分類学的な体たらくを重く見てか、命名120周年を記念して、カーによるCM 9380の頭骨要素の詳細な――1906年以来となるまとまった再記載が今年になって出版された。CM 9380――ティラノサウルス・レックスのホロタイプの頭骨について現代的な記載を書き連ねたカーは、近年命名されたティラノサウルス属3種――ポールやダールマンらによる分類を一蹴したのである。
かくしてCM 9380はホロタイプとしての存在感を久方ぶりに発揮し、再びティラノサウルスの群れの先頭に立った。マーシュが夢想しオズボーンの野望を乗せたそれは、ブラウンの最後の忘れ物――AMNHに最後まで残っていた肋骨を組み込まれ、今日もどこ吹く風でMOR 980からエドモントサウルスを守り続けている。