
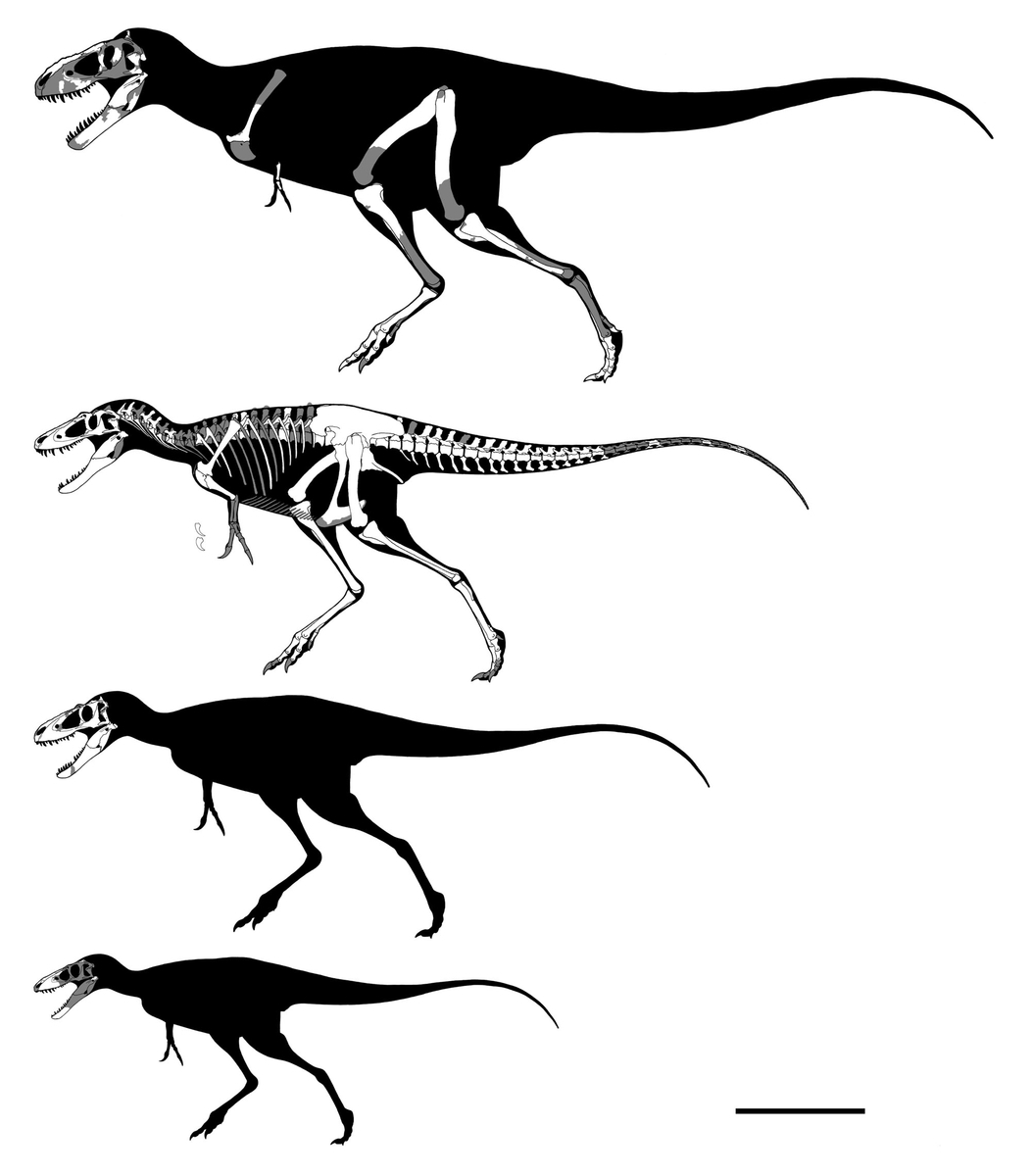
↑ティラノサウルス・レックスCM 9380(ホロタイプ;旧AMNH 973;上)と
NHMUK R7994(“ディナモサウルス・インペリオスス”のホロタイプ;
旧AMNH 5866;中)、AMNH FARB 5027(下)
スケールバーは1m。
CM 9380とAMNH 5027の全長がほぼ同じ点に注意。
CM 9380はティラノサウルスの成体としては特に後肢が長いようである。
AMNH 5027では椎骨や腰帯が異常に癒合している。
チューロニアン~コニアシアン――白亜紀の半ば過ぎに北半球で起きた恐竜相の転換によってカルノサウルス類から頂点捕食者の座を簒奪したコエルロサウルス類の一派――ティラノサウルス上科のうちアークトメタターサルを備えていたグループはその後大型化を続け、ついにカンパニアンの中ごろには全長10mを越え、カルカロドントサウルス類――それ以前の北半球における頂点捕食者に匹敵するサイズに至った。この全長10mを越える大型ティラノサウルス類――ティラノサウルス族のうち、最大最後のものこそティラノサウルス・レックスであった。
20世紀初頭、ひいては19世紀末から知られていたこの恐竜は、驚くほどの幸運――研究初期から複数の良好な骨格に恵まれ、瞬く間に6600万年ぶりの玉座――最大最強の肉食恐竜の座へと返り咲いた。様々な全長10mを超える大型獣脚類が知られるようになった今日にあってなお、ティラノサウルスは玉座から腰を上げようとはしないのである。
第一次化石戦争の“中期”――1870年代後半の主戦場はアメリカ西部へ移っていたが、その中でも目標となっていたのはもっぱらモリソン層――上部ジュラ系であった。アメリカを代表するジュラ紀後期の恐竜が軒並みこの時に発見されたことは言うまでもないのだが、その前夜――1870年代前半には上部白亜系での恐竜化石の探索も散発的に行われていた。1873年、コープはUSGSの調査隊の一員としてコロラドからいくらかの白亜紀後期の恐竜化石を持ち帰った――が、これらはハドロサウルス類と(当時まだグループとして認識されていない)角竜の残骸の寄せ集めでしかなかったのである。
やはりというか、運はマーシュに向いていた。1873年の暮れ、コロラドのデンヴァー~ゴールデン一帯で化石を採集していたベルソーという男がマーシュへ自らの発見を知らせたのである。ベルソーの言う化石にマーシュは取り立てて興味を示すこともなかったのだが(コープの調査でろくな化石が出てくる印象を持たなかったのかもしれない)、ベルソーはめげずにマーシュへ化石を送った。これはアーサー・レイクス(マーシュの下でモリソン層を掘りまくるのはもう少し後の話である)がだいぶ前に採集してベルソーへ贈ったもので、今日でもYPM 4192として残っている。この標本こそ、最初のティラノサウルス・レックス――少なくとも最上部白亜系のティラノサウルス科の化石としては初めて発見されたものであった。
ベルソーは結局マーシュの気を引くことをあきらめ、地元の地質調査にあたっていた面々――デンヴァーで教師兼地質学者をしていたキャノンやUSGSでこの一帯の地質図の作成に取り組んでいたクロスと密接に協力するようになった。ベルソーの化石は紛れもない恐竜であり、北米西部に広がる「褐炭層(今日の様々な陸成の上部白亜系の総称)」の時代を決定するのに重要だったのである。レイクスやベルソーが化石採集を行っていた(先述のティラノサウルスらしき歯の産地でもある)テーブルマウンテンの斜面ではその後大型肉食恐竜の顎(恐らくはティラノサウルス類であろう)なども発見されたが、これは現存していない。
結局、北米西部の最上部白亜系がマーシュの興味を引くまでにはそこから10年以上の時間を要した。キャノンが発見した「デンヴァーバイソン」、そしてハッチャーのトリケラトプスとの出会いを経て、マーシュは北米西部の「ケラトプス層」あるいはララミー層――今日のランス層やデンヴァー層、そしてヘル・クリーク層に目を向けたのである。悲しいかなコープにはもはやそちらへ対抗手段を送る力は残っていなかった。
かくしてハッチャーは数年にわたってランス層に居座り、数十体分におよぶトリケラトプスやトロサウルスの化石をYPMへ送りつけた。マーシュはせっせとそれらを記載してハッチャーの労に報いたが、当然ハッチャーが送ってくるのは角竜だけではなかった。
1890年、マーシュはトリケラトプスの新種――トリケラトプス・プロルススと共に、鳥に似たつくりの足を持つ小型の恐竜――オルニトミムス・ヴェロックスOrnithomimus veloxを命名した。これは「コロラドのケラトプス層」(=デンヴァー層)のもの(中手骨と足しか残っていなかった)であったが、マーシュは同時に「モンタナの同時代層」(実際にはデンヴァー層よりもだいぶ古かった)から、より大型のオルニトミムス属も命名した。オルニトミムス・ヴェロックスに倍するオルニトミムス・テヌイスO. tenuisに、3倍はあろうかというオルニトミムス・グランディスO. grandisである。マーシュはこの「第III中足骨の近位端が縮小し、第II・第IV中足骨の近位端が背側面で接する」恐竜に、新属だけでなくオルニトミムス科という新たな分類を与えた。第III中足骨の近位側が顕著に細くなり、また長骨が中空化するという鳥類的な特徴を備えた恐竜にはふさわしい名前のように思われた。
ハッチャーがせっせとトリケラトプスを掘り当てていた「ワイオミングのケラトプス層」(=ランス層)でも、オルニトミムス・グランディスらしき部分骨格が続々と発見された。これらの中には大腿骨の長さが1mを超えるもの(USNM 6183)があったが、これは当時知られていた獣脚類の中でも断トツで最大のものであることをマーシュは見て取った。マーシュはこの「巨大な肉食恐竜」の頭骨や他の部位――「角竜の特異な防御装備に対するべく、攻撃のための特別な特殊化を遂げているであろう」部位の発見を待ちわびたが、結局目にすることのないまま死んだ。
(オルニトミムス・テヌイスや、オルニトミムス・グランディスとされた様々な時代の化石は、今日いずれも(様々な)ティラノサウルス類とみなされている。ランス層産のものはティラノサウルス・レックスとみて間違いないのだが、ランス層ではいまだにまともなティラノサウルスの骨格の産出がない(後述のAMNH 5866が最良のものである)。ハッチャーをもってして首なしの部分骨格の発見に留まったのも無理はない話であり、このあたりの様子はヘル・クリーク層とは明らかに異なる。オルニトミムス科の実態が明らかになるのは第二次化石戦争を待たねばならない。)
ハッチャーの奮闘で潤うマーシュを横目に、1892年にコープもララミー層(この場合ケラトプス層と同義;マーシュが非公式名称である「ケラトプス層」を多用する一方、当然のことながらコープがこの名称を使うことはなかった)の“アガタウマス科”(言うまでもなくケラトプス科と同義)の記載に乗り出した。マーシュに先んじて各地のララミー層に唾を付けていたはずのコープであったが、この時期は腕利きの化石ハンターを欠いていたこともあり、完全に出遅れたのである。コープの手持ちはハッチャーを擁するマーシュと比べてだいぶ寂しい状況で(7m超級と思しき“アガタウマス”の部分骨格はあったが。そのくらいがやっとであった;この標本が現在どうなっているのかは不明)、コープは苦しい手持ちの中からいくつかアガタウマス科の新種をひねくりだした。そのうち新属新種としたのがAMNH 3982――2つの椎体(うち1点は恐らくコープの死のどさくさで行方不明)に基づくマノスポンディルス・ギガスManospondylus gigasであった。コープはこの巨大かつ含気化の進んだスポンジ状の2つの「胴椎」(例によって図も何もなかった;現存するものは今日第10頸椎とみなされている)を雰囲気でハドロサウルス類ではなく角竜――アガタウマス科とし、「含気孔」が存在する点で他のいかなるアガタウマス類とも異なるとしたのであった。コープの所見では、この恐竜はララミー層ではアガタウマスをしのいで最大の動物のはずだった。
コープは1897年に、マーシュは1899年に相次いで死に、残されたコープのコレクション(ANSPに収められていなかったもの)がAMNHへ流れる一方、マーシュのコレクションはYPMとUSNMへ収まった。そして世紀が変わると、ハッチャーは今やマーシュの跡を継ぐ角竜のスペシャリストとして、AMNHに入ったコープの角竜の整理を(カーネギー博物館から出張して)任される立場になっていた。
コープの論文の大半が図を欠いていた(標本番号も示されていなかった)ため、コープの記載と標本を突き合わせる作業は地獄と化した。ぼろぼろのハッチャーはAMNH 3982のナンバーを振られた椎骨――AMNHに所蔵されていたのは椎骨ひとつだけだった――を手に取り、それが角竜ではありえないことを見て取った。この椎骨はカーネギー博物館所蔵の「ワイオミングのララミー層産ドリプトサウルス」(恐らくは1902年採集のCM 1400;後述;この時期のドリプトサウルスは事実上アルバートサウルスなど真正のティラノサウルス科も含んでいる)と酷似していることに気が付いたのである(ハッチャーは1893年にYPMでの職を辞した後、プリンストン大での勤務やパタゴニアでのAMNHとの共同調査を経て1900年からカーネギー博物館に勤めていた)。ハッチャーは執筆中の角竜のモノグラフの中でマノスポンディルスが大型獣脚類であることを記したのだが、このモノグラフが出版されることになるのはだいぶ後――ハッチャーの死後3年の過ぎた1907年になってからであった。
コープの弟子筋でありAMNHの古脊椎動物部長となっていたオズボーンの下へ、その男――バーナム・ブラウンがやってきたのは1897年のことであった。カンザス大でエルマー・リッグズとともに学んだブラウンは、在学中にウィリストン――マーシュの部下としてナイオブララ・チョークの様々な陸海空の動物化石に携わってきた男の助手としてすでに十分な経験を積んでおり、「目で見る前に恐竜の匂いを嗅ぎつける」才を買われて卒業と共にAMNHに雇われたのである。就職の翌年にはパタゴニアでの18ヶ月に及ぶ調査(業務命令を受けてわずか2時間で出発したという伝説がある)に旅立ち、ハッチャーの下で様々な薫陶を受けることになったのである。
パタゴニアから戻ってそう間を置かずにブラウンは再び調査へ旅立った。今度の調査地はワイオミングのララミー層で、ブラウンはそこで巨大な獣脚類の部分骨格と出くわした。これは頭骨の要素――それまで知られていたいかなる獣脚類よりも巨大な歯骨を含んでおり、首から後ろの要素もある程度残されていた(頸椎から前方胴椎は関節しており、腰帯も大部分が残っていた)。そして何より、骨格の周囲にはおびただしい数の皮骨が――キールのついた丸く平べったい骨板が散らばっていたのである。
1900年の時点でこれほどの化石はララミー層からはまったく知られていなかった。ハッチャーをして“オルニトミムス・グランディス”の後肢を採集するのがやっとだった現ランス層にあって、ブラウンはいきなりハッチャー越えの成果を出して見せたのである。ハッチャーの“オルニトミムス・グランディス”USNM 6183とブラウンのこの謎の獣脚類AMNH 5866は実のところ重複する部位――大腿骨を保存していたのだが、AMNH 5866のクリーニングが難航したこともあり、比較の機会は訪れなかったらしかった。AMNH 5866の母岩はやわらかかったが、一方で化石そのものも相当にもろく、勢い任せでクリーニングできる代物ではなかったのである。
メディア戦略の得意な(このあたり、かなり現代的なセンスの持ち主ではあろう)オズボーンをしてAMNH 5866は華々しくデビューを飾らせるつもりだったのだろうが、記載の準備をしている間にそれどころではなくなってしまった。1901年の秋になり、ブロンクス動物園の園長であるホーナデイ(先駆的な業績で知られる一方、人種差別で悪名高いのはオズボーンと同じであった)が、知人のシーバーが彼の牧場の近くでトリケラトプスの骨格を発見したという話をオズボーンに伝えたのである。AMNHにはまだトリケラトプスの良好な標本がなかったということもあり、この話を聞いて発奮したオズボーンはブラウンとラル(YPMでのキャリアが有名だが、この頃はまだAMNH勤めだった)を現地に送り込むことにした。
ホーナデイからの手紙を頼りに、ブラウンとラルが現地に到着したのは1902年の6月になってからだった。すでに牧場の経営者はシーバーではなくなっていたが、二人は首尾よく見事な“ステルロロフス”の頭骨(AMNH 970;トリケラトプス・セラトゥスとして記載・展示されたが、これはトリケラトプス・ホリドゥスとみて間違いないようだ)を採集した。これは吻の一部と角を欠くとはいえ、頭骨の推定サイズは2mを超えるうえに保存もよかった(しかもかなり完全な前肢が残っており、後にAMNHがトリケラトプスの復元骨格を制作する際に参考となった)のだが、ブラウンとラルはもうひとつ、「マーシュが記載していない」大型獣脚類に出くわしていた。
とんでもないものを発見したのは発掘中から明らかなことであった。最初に見つかった大腿骨はざっと4フィート2インチ――1.3m近くあり、その後も続々と巨大なパーツが(ノジュール化した砂岩に半ば埋もれて)出現したのである。発掘は1シーズンで終わるものでは到底なかったが、とりあえず掘りあがったものから(最寄りの鉄道駅までの160km近くを馬ぞりで運びつつ)AMNHへ送られることになった。
ブラウンが現地に残っていつ果てるとも知れない発掘を続ける一方、(作業手当が付きたために)先にAMNHへ戻っていたラルがクリーニングを主導することになった。ブラウンによる発掘は1905年いっぱいまで続き、ミラーとカイゼンまで加わったクリーニング作業がそれに続いた。この頃オズボーンの耳にもCM 1400――1902年にアターバック(YPM時代のハッチャーの助手)がワイオミングのララミー層で採集した大型獣脚類の報が入り、思うように進まないブラウンの骨格――AMNH 973のクリーニングに焦燥感を募らせていった。AMNH 973は頭骨のかなりの部位を保存しているようだったが、一方でCM 1400にも頭骨の要素が相当に含まれているという話だったのである。両者は同じ種を代表している可能性があり、そうだとすればもはや早い者勝ちの状況であった。
1905年10月、オズボーンはAMNH 973とAMNH 5866の記載に踏み切った。AMNH 973は発掘すら終わっていなかったが、それでも相当な部位が残っているのは間違いなく、とにかく先手を打って命名だけしておけばなんとかなると見てのことであった。かくしてここに、ティラノサウルス・レックスTyrannosaurus rexが命名されたのである。急ごしらえの命名であった(AMNHに到着していた部位でさえクリーニングが終わっていなかったため、ひたすらに巨大である以上のことはオズボーンにも言えず、体サイズの割にやたら小さい上腕骨がこの動物に属することさえ断言できなかった)が、それでもオズボーンはこの巨大な肉食恐竜にとてつもない可能性を見出していた。
オズボーンをもってして、ひたすらにデカいこと、頭骨に粗面からなる装飾が発達していること、そして皮骨板を持たないらしいこと程度しか述べられない状態(保存されていることが確認できた部位でもってざっくりとマシューの描いた骨格図を掲載してもいるのだが;研究の「総決算」として骨格図が描かれるかどうかであったこの時代に、ざっくりでいいからとにかくビジュアルを、というオズボーンのセンスはやはり先駆的であった)のティラノサウルスに対し、次のページで命名されたディナモサウルス・インペリオススDynamosaurus imperiosus――AMNH 5866のクリーニングは終わっており(AMNH 973に比べればだいぶ不完全な骨格ではあった)、背中を鎧で固めた巨大肉食恐竜の姿は明らかなように思われた。オズボーンはディナモサウルスの見事な歯骨と歯について詳しく記載することができたのである(ティラノサウルスでも歯骨が保存されているのは間違いなかったが、クリーニング中のためにディナモサウルスとの比較は全く行えなかった)。最後にオズボーンはカナダ産ドリプトサウルスのために新属新種――アルバートサウルス・サルコファグスを命名し、論文の締めくくりとした。
AMNH 973のクリーニングは1906年に終わり、ようやくティラノサウルス・レックスの実態が明らかとなった。早速オズボーンはティラノサウルスについての第2報を出し、これについて詳しく記載を行った(ついでにティラノサウルス科も設立した)――が、結果は意外なものとなった。AMNH 973とAMNH 5866は形態的に区別できそうな点が見当たらなかったのである。カーネギー博物館に対しどうにか先手を取ったオズボーンだったが、ディナモサウルス・インペリオススをティラノサウルス・レックスのシノニムとすることでそのツケを払うことになったのだった(すんなり自分のミスを認めているあたり、まるっきり同じ形をしていることによほど驚いたのかもしれない)。ディナモサウルスの方が標本の発見では先んじていたものの、論文内での命名順が仇になったのである。
この時点でオズボーンの手元にはクリーニングの終わった3体目のティラノサウルスの部分骨格AMNH 5881があり、3体を合計すると尾を除く全身の相当な部位のパーツが揃った計算であった(尾の要素はAMNH 973に尾椎の残骸が1点含まれていた程度であった)。AMNHの誇るサイエンスアーティストの一人であったスターリングはこれら3体を取り混ぜ、見事な骨格図を描いてみせたのである。
原記載でどうするかさんざん悩んだ末に骨格図へ描き入れられていたAMNH 973の小さな上腕骨は、結局AMNH 973そのものと判断された(スターリングの骨格図でも、マシューによる原記載の図と同様3本指の小さな手が復元されている)。ひどく退縮しているとはいえ筋肉の付着点は明確に残されており、なにがしかの機能――おそらくは交尾の際に背中につかまるのに役立ったのだろうとオズボーンは考えた。
AMNH 5866――ディナモサウルスのホロタイプに含まれていた皮骨の解釈はもっとずっと厄介であった。AMNH 973やAMNH 5881で皮骨はひとかけらも見つからなかった一方で、AMNH 5866からは相当数の皮骨が発見されたのである。AMNH 5866が他の様々な恐竜とごった煮になっていたのも確かであった――トリケラトプスと思しきフリルの断片や、ハドロサウルス類の顎の断片も共産したのである。一方で、AMNH 5866は多数の肋骨――紛れもない獣脚類の形態のそれも保存しており、胴回りのパーツがかなり残っている格好でもあった。かくしてオズボーンは、ティラノサウルスの背中が皮骨で装甲されていたと考えることにしたのである。これほどのサイズの肉食動物が身を守るための装甲を発達させるのも妙な話ではあったが、オズボーンはティラノサウルス科――ことによるとティラノサウルス本人から身を守るためのものと捉えたのであった。
(AMNH 5866の皮骨はアンキロサウルスのものと考えられるようになって久しい。ブラウンは1908年のアンキロサウルスの原記載の中で「ティラノサウルスの皮骨」とアンキロサウルスのそれを比較しているが、形態の違いから両者は別物と判断している。)
1906年の暮れ、AMNH 973の巨大な腰帯と後肢が組み上げられ、化石ホールに展示された。大きな鳥の骨格が足の間にちょこんと収まり、ティラノサウルスの途方もないサイズとそこはかとない鳥との類似性を訴えかけたのだった。ナイトによって描かれた復元画が花を添え、命名から1年ほどのうちに史上最大最強の陸生捕食者のイメージが完成したのである。
(AMNH 973の頭骨も(当時の常識として、実物を石膏に練り込む形で)復元されたが、これは今日の目で見るとだいぶ不正確な代物であった。未産出の部位はアルバートサウルスの頭骨でも補完できなかったのである。売却先のカーネギー博物館でAMNH 973がマウントされた際にも、頭部に据えられたのはこの復元頭骨(のレプリカ)であった。全身のマウントのために椎骨のアーティファクトの制作も進められたが、これをAMNHで展示することはとうとうかなわなかった。)
1908年、ブラウンは再びモンタナのララミー層を訪れた。1906年の予備調査で発見したハドロサウルス類やらなんやらの発掘が当面の目標だったのだが、ふたを開けてみればこれはハズレであり、結局発掘をあきらめることにした。が、キャンプへの道すがら、ブラウンは生涯最高ともいえる化石との出会いを果たしていた。4つの風化した尾椎が露出しており、ちょっと掘ってみただけで15個の関節した尾椎――角竜ともハドロサウルス類とも異なっており、ブラウンのまだ見たことのないタイプの尾椎だった――が姿を現したのである。その根元は丘へと続いており、腹をくくったブラウンはキャンプを骨格のそばへ移動させた。
この標本――ブラウンは“バグ”と呼んだ――の発掘は1908年の7月から始まり、すぐにそのすさまじい様相が明白となった。ダイナマイトを駆使して丘の上部を吹き飛ばすのに1週間を要したが、そこにあったのは体の右側を下にして横たわったティラノサウルスの巨大な死体――尾の中ほどから首の先までデスポーズで関節した骨格であった。右側の肋骨は全て関節状態でそこにあり、左側の要素も全てがそのあたりに散らばっていた。前後肢はそっくりなくなっていたが、恥骨と座骨の間に引っかかっていた頭骨は完全な状態――頭蓋も下顎もすべて揃っていたのである。大量の堆積物で覆われていたぶん(=風化から相当に保護されていた)化石の保存はよく、AMNH 973の時のように意味不明に硬い砂岩のノジュールに埋まっているということもなかったので、化石の層準に達してしまえば掘り出すのはそう難しい作業ではなかった。9月の上旬には発掘は完了したのである。
ニューヨークへ到着した“バグ”は、AMNH 5027のナンバーを与えられ、さっそくクリーニングと展示準備が始まった。尾の後半を除けば体軸の要素は完全な状態であり、そしてこの標本はもうひとつ奇跡的な幸運に恵まれていた――AMNH 973と事実上同サイズの動物だったのである。AMNH 973は肩甲骨と上腕骨、それにほぼ完全な後肢を保存しており、AMNH 5027と相互補完することで下腕と尾の後半を除くほぼ全身のパーツが揃ってしまう格好であった。プレパレーションにはAMNHの技術の粋が注ぎ込まれ、AMNH 973のキャストで補完しつつ未知の部位はアロサウルスを参考としたマウントの準備が整ったのである(AMNH 973の第III中足骨は遠位端しか残っておらず、制作されたアーティファクトの足は非アークトメタターサルであった。これは数年後に明らかな誤りであったことが確認された(後述)が、いかんせんマウントの根幹をなす部分でもあり、前肢と違って交換することはかなわなかった。趾骨のアーティファクトは(末節骨がだいぶ大きいものの)特別問題のあるものでもなく、リノベーションの際も中足骨ごとそのまま再利用された)。
せっかく同サイズのティラノサウルスが2体もある以上、オズボーン(1908年に館長に就任していた)としてはAMNH 973とAMNH 5027を並べて展示したいところであった。それも単に2体を並べるのではなく、生息時のようにいきいきとして「映え」るポーズで、である。AMNH 5027の頭骨の記載を済ませたオズボーンは、クリストマン――スターリングと双璧をなすAMNHのサイエンスアーティストであった――に1/6スケールでティラノサウルスの骨格模型を2体制作させ、マシューと二人でマウントのポージングと配置を検討した(ポーズ案①、②)。最終的に決定されたポーズ案はブロンクス動物園の爬虫類部門のスタッフの意見を参考としたもので、AMNH 973が仕留めた“トラコドン”をAMNH 5027が奪おうとするものだった。“トラコドン”に覆いかぶさるようにして咆哮するAMNH 973と、威嚇のために限界(以上)まで背中を起こしたAMNH 5027――頭のてっぺんは高さ5.5mに達した――というあまりにもダイナミックな取り合わせはしかし、様々な意味で無理筋であった。重い化石と石膏の組み合わせをこれほどアクティブなポーズで組み立てた前例はなく(今日でもない)、今日の目で見ても抜きんでた技術を誇っていた(化石にはあまりやさしくなかったが)AMNHの精鋭プレパレーターたちをもってしても、これほどのマウントを実際に組み立てられる保証はなかった。そもそも展示ホール(当時、脊椎動物化石は全て今日の「脊椎動物の起源」ホールに押し込められていた;1921年になって現在の「鳥盤類ホール」が完成し、恐竜の展示は全てそちらへ移ったが、それでも決して余裕のある空間ではなかった)のサイズからしてこの巨大な動物2体を対決させるのは不可能だったのである。そしてこのあたりの問題をクリア――ホールの増築をするためには不可欠である資金がどうしようもなかった(このあたりはオズボーンをオズボーン足らしめる得意分野ではあったのだが、それでも限界はあった)。
結局、AMNH 5027をメインにしたマウントただ1体が1915年にデビューすることになったが、この恐竜――ティラノサウルス・レックスを真に恐竜の代表とするにはそれで十分であった。この「世界で唯一のティラノサウルスの復元骨格」は公開されるやいなやセンセーションを巻き起こし、その後30年近くに渡って世界で唯一というその立場を守り続けたのである。1917年にオズボーンはAMNH 5027の椎骨や腰帯についても記載・図示を行い、ティラノサウルスに関する研究のひとまずの締めくくりとしたのであった。
(AMNH 5027のマウントが展示されてからしばらくのうちは、1906年に完成したAMNH 973の後肢のマウントもすぐ脇に展示され続けていた。AMNH 5027のマウントは台座ごと移動させることが可能であり、上述の展示室移転時には大いに役立ったようである。オズボーンは1917年の論文の中でマノスポンディルスに関するハッチャーの意見を追認し、さらにティラノサウルスの後方頸椎~第1胴椎と酷似していることを述べた。が、マノスポンディルスが椎体2個のみに基づくことから、これを属種不定のティラノサウルス類とした。オズボーンはさらに、当時スミソニアンでマウントされたばかりであったケラトサウルスのホロタイプの皮骨について触れ、“ディナモサウルス”の皮骨がティラノサウルスに属することはほぼ確実と考えていた。ゴルゴサウルスの発見によってランベはティラノサウルスが2本指であった可能性を指摘するようになり、これを受けて1927年にAMNH 5027の前肢は交換された。1920年にギルモアがスミソニアンの所蔵する様々な獣脚類を一気に記載した際にハッチャーの採集した一連の“オルニトミムス・グランディス”がティラノサウルスであることが指摘され、マシューとブラウンも1923年にこれを追認した。ここに至ってティラノサウルスがアークトメタターサルを持っていたことがようやく認識されたのである。
マノスポンディルス・ギガスがティラノサウルス・レックスのシニアシノニム云々の問題は(目を引く話題ゆえに)相当な誤解を持って語られがちであるが、オズボーンから今日に至るまできちんとした比較でもってマノスポンディルスがティラノサウルスであると断定した研究者はおらず(恐らくはティラノサウルスで間違いないのだが)、ラーソンがマノスポンディルスの掘り残しの可能性があるとして喧伝したBHI 6248(これはまぎれもないティラノサウルス・レックスで、“E. D. コープ”の愛称が付いている)についても明らかな証拠は何もなかった。「マノスポンディルス・ギガスがティラノサウルス・レックスで間違いないとして」前者はICZN第4版(2000年以降適用)の遺失名nomen oblitumに該当し、自動的に後者が保存名nomen protectumとなるが、そもそもマノスポンディルス・ギガス(と確実に言える標本)が断片的すぎて種としての特徴をなにも保持しておらず(単に巨大なティラノサウルス類の椎骨としか言えない)、保護名うんぬんの状況にさえ(おそらく永久に)至っていないのが現状である。遺失名云々も、特にティラノサウルス・レックスという学名の保護を目的として設けられた強権措置ではない。)
結局CM 1400は記載されず、ティラノサウルスを巡る情勢はオズボーンの一人勝ちに終わった――のは、オズボーンが死ぬまでの間だけであった。大恐慌の影響をまともに食らってAMNHの入館者数は減り続け、オズボーンが館長を退くと(元部長のオズボーンが館長職についていたがために)金食い虫であった古脊椎動物部門は地質部門に統合され、予算と人員は大きく削減された。こうした状況は割とどこでも似たようなものであり、かくして恐竜研究の暗黒時代が訪れたのである。
予算不足にあえぐAMNHがとった最後の手段が標本――バックヤードで眠っていたティラノサウルス・レックスのホロタイプAMNH 973の売却であった。第二次世界大戦はとうに始まり、対日戦の足音も近い中にあってニューヨークは決して安全ではなかった――が、交渉はそのさらに以前から始まっていた。1941年、「戦禍を避けるために」AMNH 973を買い取ったのはよりにもよってカーネギー博物館――ララミー層の大型獣脚類の命名を巡ってAMNHに先を越された格好となった――で、翌1942年にはAMNH 5027のキャストと組み合わせた復元骨格が制作されたのである。AMNH 5027のマウントと同様に実物頭骨を足元に従えたそれ――CM 9380は、ディプロドクスやアパトサウルスと正対させられたのであった。暗黒時代の中で、AMNHは「世界で唯一のティラノサウルスの復元骨格」を展示しているという立場を自ら手放したのである。
(AMNH 973とともに脳函AMNH 5029もカーネギー博物館へ売却された。こちらは今日CM 9379として所蔵されている。)
やがて暗黒時代は終わり、恐竜ルネッサンスの号令が響く1960年代後半にあってAMNHからはさらにティラノサウルスが旅立っていった。寄付金を得て勢いづく大英自然史博物館に購入されたのはAMNH 5866――“ディナモサウルス・インペリオスス”とAMNH 5881で、それぞれBMNH(現NHMUK)R7994とR7995となった(さらに“ディナモサウルスの皮骨”はR8001として分離した)。
ロンドンへやってきた“ディナモサウルス”たちをAMNH 5027のキャストと組み合わせてマウントするという任についたのはニューマンだったが、彼はこれをAMNH 5027やCM 9380のマウントと同様の古式ゆかしいスタイルで組み立てる気はなかった。恐竜ルネッサンスによってこの「ゴジラ立ち」スタイルは古ぼけつつあったし、展示空間としてもティラノサウルスの頭を目いっぱい持ち上げて組み立てる余裕がなかった(天井の高さがあまりなかった)。ニューマンはAMNH 5027のキャストの首と下顎をR7994で置き換えつつ、R7995に基づいた腹骨を添えたウォールマウントを組み上げたのだが、これはAMNH 5027とは相当に異なった見てくれになっていた。AMNH 5027では計53個として復元された尾椎のトータル数をゴルゴサウルス(の推定数)と同様の37個まで減らし、3.6mも短くなった尾を常時地表から持ち上げるものとして復元したのである。
(オズボーンはあくまでAMNH 5027を活発な動物として復元しており、愚鈍さゆえに尾を常時引きずっていたわけではなかった点に注意が必要である。1917年に詳細に記載されたゴルゴサウルスのホロタイプは尾椎の大多数を保存しており、AMNH 5027のマウントの完成から数年のうちには長すぎる尾の問題が明らかになっていた。ランベはゴルゴサウルスの尾椎のトータル数を36個前後と推定していたが、その後制作されたAMNHのゴルゴサウルスのマウントでもトータル数は37個とされたのであった。ティラノサウルス(や他のティラノサウルス科)の尾椎のトータル数は今日でも厳密にはわかっていないが、“スー”で37個以上あったのは確実であり、またタルボサウルスの大型幼体で45個あったことが確認されている。ティラノサウルスやその他のティラノサウルス科も同程度で間違いないだろう。個数としてはランベやニューマンの推定からだいぶ増えているようにも思えるが、「増加分」の尾椎のサイズはごく小さなものであり、全体で言えばランベやニューマンの復元にひょろっとしたわずかな延長分が付くに過ぎない。)
1970年、このあたりの話題についてニューマンは論文を出版した。尾の復元に関するニューマンの指摘は至極もっともであり、獣脚類の研究者の間で1920年代初頭には暗黙の了解となっていた案件を論文化しただけであったといえばそうでもあった。これも恐竜ルネッサンスのひとつだったのである。さらにニューマンは走鳥類とティラノサウルスを比較し、まともな尾を持たない前者が大腿骨をほぼ水平に保つことで足を重心(胴体の中央付近)の真下に置いている一方、(以前よりだいぶ短く復元したとはいえ依然として)長く重い尾を持つ後者の大腿骨が走鳥類のような姿勢を取らせることなどできそうにないことに注目した。そして、ティラノサウルスの膝関節はほどほどに伸びた状態(依然として曲がった状態である)が基本――足は胴と腰の境界あたりの真下に来る――だとして、長く重い尾で上半身とうまくバランスがとれることに気付いたのである。ニューマンはここに、ティラノサウルス(や同様のボディプラン(重い頭と上半身、ほどほどに長く重い尾)を持つ他の大型獣脚類)が、走鳥類と同様、通常時は上半身と尾をほぼ水平に保っていたことを断言したのであった。ここでついに「現代的」なティラノサウルスの復元が現れたのである。
ニューマンのティラノサウルスはやや上半身を持ち上げ、ゆったりと行儀よく歩くように描かれていたが、同年にラッセルが出版したカナダ産ティラノサウルスのモノグラフに掲載されていたダスプレトサウルスの骨格図はずっと過激であった。上半身をやや下げて大股で獲物を捜し歩くラッセルのダスプレトサウルスは、恐竜ルネッサンスにおける大型獣脚類のイメージを決定付けるものであったのである。続いてラッセルとキッシュのコンビで描かれた様々なカナダの恐竜の復元――骨格図そして復元骨格と同じポーズで描かれたダスプレトサウルスに加え、砂埃を巻き上げてエドモントサウルスをずたずたに引き裂くティラノサウルスを含んでいた――はこれを加速させ、恐竜研究の黎明期から連綿と続いていた(そして「暗黒時代」に一度は途絶えかけた)「活発な恐竜」のイメージはここで一つの完成を見たのである。
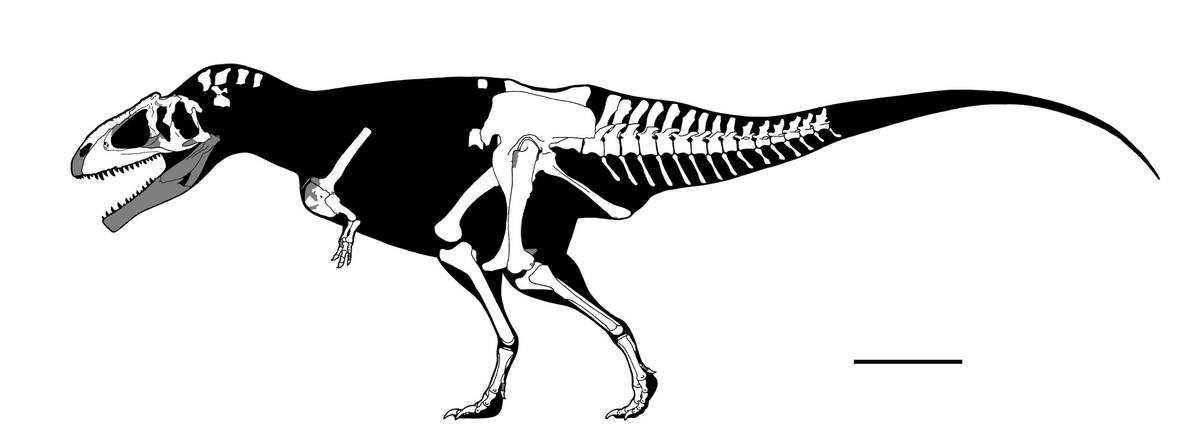
↑“ランス期”の様々な未成熟のティラノサウルス類。いずれも今日ではティラノサウルス・レックスとみなされている。
上から、LACM 23845、BMRP 2002.4.1(“ジェーン”)、CMNH 7541、LACM 28471。
スケールバーは1m。
ニューマンの意見が即座に受け入れられたわけではなかった――「半ゴジラ立ち」で復元しようとする流れはその後もしばらく続いてはいた――が、それでもこの復元は着実に根付き、いつしか「ゴジラ立ち」は時代に置いてけぼりにされた。
これにさかのぼる1967年、ロサンゼルス郡立博物館(現ロサンゼルス自然史博物館;略号はLACMのまま今日に至っている)はAMNH以来となる大規模な遠征調査を3シーズンにわたってモンタナのヘル・クリーク層――1929年にブラウンによって分離・命名されていた――で行うこととした。この遠征でLACM隊のガイドとなったのが完全独学のアマチュア化石ハンター――ハーレイ・ガルバーニであった。
ガルバーニはLACM隊を次から次へと化石の産地へ導いた。ブラウン以来の才の持ち主であったガルバーニによってLACMは有数のヘル・クリークの化石コレクションを築き上げることになったのだが、その中でもとりわけ重要な発見が3体の獣脚類――LACM 28471とLACM 23844、そしてLACM 23845であった。
LACM 28471――もっぱら“ジョーダンの獣脚類”の通称で呼ばれた――は遠征に先立つ1966年に採集されたもので、これは奇妙な小型獣脚類の頭骨の残骸であった。風化した上下の吻の要素と後頭部の断片が残されており、もともとはティラノサウルスの幼体と漠然と考えられていた。一方、これの研究にあたったモルナーは“ジョーダンの獣脚類”がティラノサウルス科と多数の特徴(例えば「D字断面の前上顎骨歯」;LACM 28471は前上顎骨を欠いていたのだが、一緒に見つかった歯のうちのひとつは前上顎骨歯と判断された。これは鋸歯を欠いていた)を共有する一方で、頭蓋天井の縫合線がだいぶ異なるらしいことも見て取った。“ジョーダンの獣脚類”の吻はやたら低いらしく、ティラノサウルスは言うまでもないとして、ヘル・クリーク層産のもう一つのティラノサウルス類――アルバートサウルス・ランセンシスと比べても相当に低かったのである。“ジョーダンの獣脚類”はドロマエオサウルス類とも相当数の特徴を共有しているらしく、1978年にモルナーはひとまず大型のドロマエオサウルス類とみなした。
続く1981年、モルナーはLACM 23845――遠征調査中にエンダールの牧場で採集された中型のティラノサウルス類を記載した。これは巨大なティラノサウルス・レックスLACM 23844――ばらけた頭骨の大部分に加えてほぼ完全な足が保存されていた――の発掘のさなかに発見されたものであった。エンダールは自らブルドーザーを駆って発掘に協力したのだが、その排土の山の中に(ブルドーザーによって)ぼろぼろになったティラノサウルス類の骨格が入っていることにガルバーニが目ざとく気付いたのである。産状が産状だったためにこのティラノサウルス類には複数の標本番号が振られていたが、それらはやがて同一個体であるとみなされてLACM 23845に統一された。これは頭骨の様々な部位の断片(下顎はそれなりに完全だった)と後肢の大部分、部分的な肩帯と前肢を保存していたのである。。
この骨格も当初ティラノサウルスの未成熟個体とされていたのだが、その後cf. アルバートサウルス・ランセンシスとされていた。モルナーもやはりLACM 23845はアルバートサウルス(この場合ゴルゴサウルスを含む;この当時、ドリプトサウルスもアルバートサウルスに近いものであるらしいと考えられるようになっていた)的な代物であると考えた。ロジェストヴェンスキーによって、ティラノサウルス類の亜成体が成体と比べてずっとほっそりしていることが指摘されており、これはLACM 23845にも言えそうであった(頭蓋天井の結合は相当緩かった)が、一方で頭蓋天井の縫合線のパターンはティラノサウルス・レックスとは別物で、アルバートサウルス的な(後頭部の幅が左右方向に急拡大しない)頭骨のつくりであることは間違いなかったのである。脛骨の近位端の形態はドリプトサウルスとよく似ているように思われた――が、腓骨の形態は全く異なっており、モルナーはLACM 23845がティラノサウルスの亜成体でもドリプトサウルスでもないと判断した。残るマーストリヒチアンの北米産ティラノサウルス類といえばアルバートサウルス属とダスプレトサウルス・トロススだけで、ヘル・クリーク層といえば上述の通りアルバートサウルス・ランセンシスの産地でもある。アルバートサウルス・ランセンシスの頭蓋天井はしっかりと結合しており(実際はまったくそんなことはなかったのだが)、LACM 23845のゆるいそれとは異なるようにも思われた――が、同じ地層から産出しているということから、ひとまずモルナーはLACM 23845をアルバートサウルス・cf. ランセンシスとしたのであった。
(ゴルゴサウルス・ランセンシスGorgosaurus lancensis――ラッセルによる1970年のモノグラフ以来アルバートサウルス・ランセンシスと呼ばれていた――は、1942年に「モンタナのランス層」でクリーブランド自然史博物館のダンクル(ダンクルオステウスは彼への献名である)によって採集された頭骨に基づいている。この頭骨CMNH 7541はほぼ完全な一方で相当に変形・破損していたが、各部の縫合線はかなり強固に“癒着”しているように思われた。このためこれは成体とみなされたのである。ロジェストヴェンスキーはこの標本が全体的な形態からしてティラノサウルスの幼体に過ぎないことを指摘したのだが、一方でラッセルは縫合線の様子からして成体で間違いないとみなした。LACM 23845はCMNH 7541と比べてだいぶ大きな個体ではあった一方で未成熟個体であることも間違いなく、モルナーはLACM 23845をアルバートサウルスcf. ランセンシスとはしたものの、再三にわたって生層序的な考え方に基づくもの(=直接的な形態の類似に基づくわけではない)ことを記している。)
ヘル・クリーク層の獣脚類に関するモルナーの慎重な意見を吹き飛ばしたのはポールであった。ポールは1988年に出版した肉食恐竜事典の中で、LACM 28471の「鋸歯を欠いたD字断面の前上顎骨歯」がアウブリソドンに他ならないことを指摘し、LACM 28471がアウブリソドン類であることを指摘したのである。模式種であるアウブリソドン・ミランドゥスがずっと古いジュディス・リバー層からの産出であることから、ポールはこれにモルナーにちなんでアウブリソドン・モルナリスAublysodon molnaris(末尾のsが余計である)の名を与えた。モルナーとカーペンターは1989年にLACM 28471を再記載してポールの意見を追認しつつ、前上顎骨歯が形態的に区別できなかったことから、種小名についてはcf. ミランドゥスに留めた。
アルバートサウルス・ランセンシス――CMNH 7541は、1988年にバッカーとウィリアムズ、そしてカリーによってティラノサウルス科の小型の新属――ナノティラヌスNanotyrannus(所蔵先にちなみ、もともとはClevelanotyrannusという属名を考えていたらしい)に移された。進化的な特徴――両眼視のために著しく後頭部の左右幅が拡大しており、かつ極端に左右幅の狭い吻を持っているように思われた――と原始的な特徴――歯の数はだいぶ多かった――の組み合わせからしてアルバートサウルスやゴルゴサウルスと同じ属のようには思われなかったのである。バッカーらはナノティラヌスがティラノサウルス科の中でもっとも原始的なものとみなし、進化的な特徴はティラノサウルスへの収斂とみなした。
ポールはこれを受け、LACM 23845をアルバートサウルス属の新種――アルバートサウルス・メガグラキリスA. megagracilisとした。ここにヘル・クリーク層の4つのティラノサウルス類――大きい順にティラノサウルス・レックス、アルバートサウルス・メガグラキリス、ナノティラヌス・ランセンシスそしてアウブリソドン・モルナーリが揃ったのである。オルシェフスキーは後にLACM 23845がアルバートサウルスではなくもっとティラノサウルスに近いものと考えてこれに新属名ディノティラヌスDinotyrannusを、LACM 28471を歯のみに基づく属であるアウブリソドンから外し、新属名スティギヴェナトルStygivenatorを与えたのだった。
1992年に出版されたカーペンターによるティラノサウルス科のレビュー(ナノティラヌスの論文や肉食恐竜事典は引用されておらず、原稿の完成は明らかに1988年以前である。もっとも、カーペンターの弁はそのあたりへの批判としても有効であった;脚注でカーペンターの所属先が最近変わったことだけが書き添えられている)は、このあたりのイケイケな状況に冷や水をぶっかけるものであった(一方で、かねてよりタルボサウルスの幼体とみなすかどうかでもめていたゴルゴサウルス・ノヴォジロヴィG. novojiloviに新属マレエヴォサウルスMaleevosaurusを与えている)。CMNH 7541の基本形はアルバートサウルスやゴルゴサウルスではなくティラノサウルスのそれであり、頭蓋天井の“癒着”だけでこれを成体とみなすことに注意喚起をしたのである。LACM 23845にしても、ティラノサウルスの亜成体ではないとする根拠はごく弱い(脛骨の近位端の形態も有意な差かといえばそうでもなさそうだった)ことを指摘したのだった。
このあたりの問題――“本職”ではないポールやオルシェフスキーが引っ掻き回したことでだいぶ面倒な事態に陥っていた――に正面から突っ込んだのはカーであった。卒論、修論を通してティラノサウルス類の様々な成長段階の個体と向き合ってきたカーをして、CMNH 7541は幼体にしか見えなかったのである(幅の広い後頭部やらなんやらはどう見ても化石化の過程における変形の産物であった)。幼い顔つきであった一方でCMNH 7541の細部の特徴は何よりもまずティラノサウルス・レックスと酷似しており、それはLACM 28471にしてもLACM 23845にしても同様であった。2004年、カーはこれらヘル・クリーク層産の問題児に関する詳細な再記載を出版し、いずれもティラノサウルス・レックスの未成熟個体であること――ヘル・クリーク層ひいては“ランス期”の北米西部のティラノサウルス類がティラノサウルス・レックスただひとつであることを明示したのである。
かくしてこれらの問題児が未成熟個体であることは誰もが認めるところとなった――が、ここからが泥沼であった。CMNH 7541や、それと多数の特徴を共有する(もう少し大型の)未成熟個体BMRP 2002.4.1――“ジェーン”に見られる種々の特徴は、これが未成熟個体であることを差し引いても分類学上有意なのではないかとする意見が(事実上ピーター・ラーソンただ一人から)延々と述べられたのである。ヘル・クリーク層から時折発見される未成熟のティラノサウルス類(その大半がプライベートコレクションへ入った)の多くはCMNH 7541と同様の特徴を有している――ナノティラヌスの未成熟個体である一方、ラーソン(や他のいくらかの業者)の弁ではいくらかまぎれもないティラノサウルス・レックスの幼体も産出しているというのであった。
カーからすれば営利業者の言う観察事実はアテにならないものでもあり(公立博物館の標本と同レベルのアクセスは実際問題として不可能であり、ラーソンの言い分が今日の形態記載において通用するものかといえばそうでもなさそうな部分が割とあった)、両者の言い分はこれといって交わることはなかった。カーの言う「成長で変化する特徴」をラーソンはひたすらに「成長で変化しない」と述べるに終始したのである。ラーソンの言うところの「ティラノサウルス・レックスの幼体」が公立博物館のコレクションに収まることもなかったのだった。
(2010年にウィットマーらはCMNH 7541の詳細な再記載――高精度のCTスキャンに基づく――を行い、ティラノサウルス・レックスの成体とは異なる様々な特徴を改めて確認したが、一方でこれらが成長段階で変化するのかどうかについて判断を下すことはなかった。ウィットマーらは様々な未記載標本の検討こそがこの分類学上の問題を解決する上で不可欠であると述べるほかなく、究極的にはそれから10年以上がたった現在でもそれは同じである。“ナノティラヌス・ランセンシスの未成熟個体”とティラノサウルス・レックスの成体に見られる形態差のうち、有意かもしれない違いは2点――歯の本数と方形頬骨の外側面にある小さな含気孔に終始する。タルボサウルス・バタール(ティラノサウルス・レックスとごく近縁とされており、カーは常々、原記載と同様にティラノサウルス属として扱っている)の幼体MPC-D 107/7ではどちらの特徴も成体と同様であったが、ティラノサウルス・レックスで同じパターンだったかどうかは判断する術がない。)
1980年代初頭、開館を控えて活発な調査を行っていたロイヤル・ティレル博物館によってカナダ――チャールズ・モートラムによるわずかな採集例だけがあった――で新発見が相次いだ。カリーらが目星をつけたのはCMN 9950――1946年にチャールズ・モートラムがスコラード層の急崖の中腹で発見し、1960年にラングストンが持ち帰ったその人――趾骨でしかなかった――であった。カリーは1980年に現場を訪れ、チャールズ・モートラムやラングストンが目にしなかったもの――侵食によって姿を現しつつある骨格を見て取り、1981年に発掘が始まった。この“ハクスリー・レックス”――TMP 1981.012. 0001は首から後ろのパーツがわりあいに揃っており、後肢がほぼ完全なだけでなく、椎骨も関節状態で残されていた。発掘隊は崖の上半分を掘り崩しつつ台地の上にクレーンを据えて発掘を行ったが、これによって発掘現場の背後には高さ30m以上に達する新たな崖が生まれた。この時点で頭骨要素は後眼窩骨1点のみであり、まだ相当なパーツが埋まっているようにも思われた――が、崖を丸ごと取り除くだけの予算はなく、これほどの崖をそびえたたせたまま狸掘りをするわけにもいかなかった。発掘はここで中断され、採集されたパーツは(チャールズ・モートラムが発掘をあきらめた程度には硬い母岩だったが)急ピッチでプレパレーションが進められた(プレパレーター6名がかかりきりで丸3年を要したという)。AMNH 5027の頭骨を据えたマウントはどうにか開館に間に合い、以来ロイヤル・ティレルのシンボル展示のようになっている。トンネルを掘って掘り残しをどうにかしようという計画も進められていたが、1993年に崖が崩落して発掘現場は完全に埋まり、かくして頓挫したのであった。
頭骨に関して言えば不完全燃焼に終わってしまったこの調査の裏で、カリーたちはもうひとつのアルバータ産ティラノサウルスにめぐりあっていた。1980年の夏に釣りに来ていた高校生が発見したそれは、暁新統――新生代の地層とされていたウィロー・クリークWillow Creek層の崖に埋まっていたのである。この骨格が埋まっていた場所はさほど高くない崖の中腹で、幸い(TMP 81.12.1とは異なり)重機が崖下から登ってこられる高さであった。しかもこの場所――バッドランドのはずれにあり、青々としたロッキー山脈のふもととそこから注ぐクロウズネスト川がよく見えた――はカナダ太平洋鉄道の大陸横断線からさほど遠くなく、標本の搬出にも不都合がなかった。地下水の運んだマンガン分で染まった漆黒の化石――灰白色の粗粒砂岩に埋まっており、はっきりと目立った――にちなんで“ブラック・ビューティー”の愛称を与えられたそれ――TMP 1981.006.0001は部分骨格に加えて頭骨の相当な部分を保存していたが、マウントを開館に間に合わせるべくTMP 81.12.1に注力していた博物館に同時並行でこれのクリーニングを進める余裕はなかった。1985年に博物館が開館してからはフルタイムでクリーニングに従事できる人数も減り、この標本のクリーニングは遅々として進まなかった。転機となったのは1990年の日立ディノベンチャー大恐竜博で、会場内の特設ブース内で頭骨のクリーニングが実演展示されたのである。帰国した“ブラック・ビューティー”は来るべきCCDPの世界巡回展で展示すべくクリーニングのピッチが上げられ、そしてCCDPの巡回展には有名なウォールマウント――分割移動式の台座にデスポーズで埋め込まれた実骨と、キャストで組み上げた3Dの全身骨格が展示されたのだった。
(“ブラック・ビューティー”の3DマウントはCCDPの特別展で展示されたものと、それ以降に新たに制作されたものとで2タイプあり、例えば下顎後方――角骨と上角骨のアーティファクトが全く異なる。茨城県自然博物館にあるのは前者のタイプであり、福井県立恐竜博物館にあるのは後者のタイプである。ウォールマウントは巡回終了後にロイヤル・ティレルで常設展示されて今に至っているが、この時に足やらのパーツを(実骨準拠の?)ハイディテール版に交換されている。)

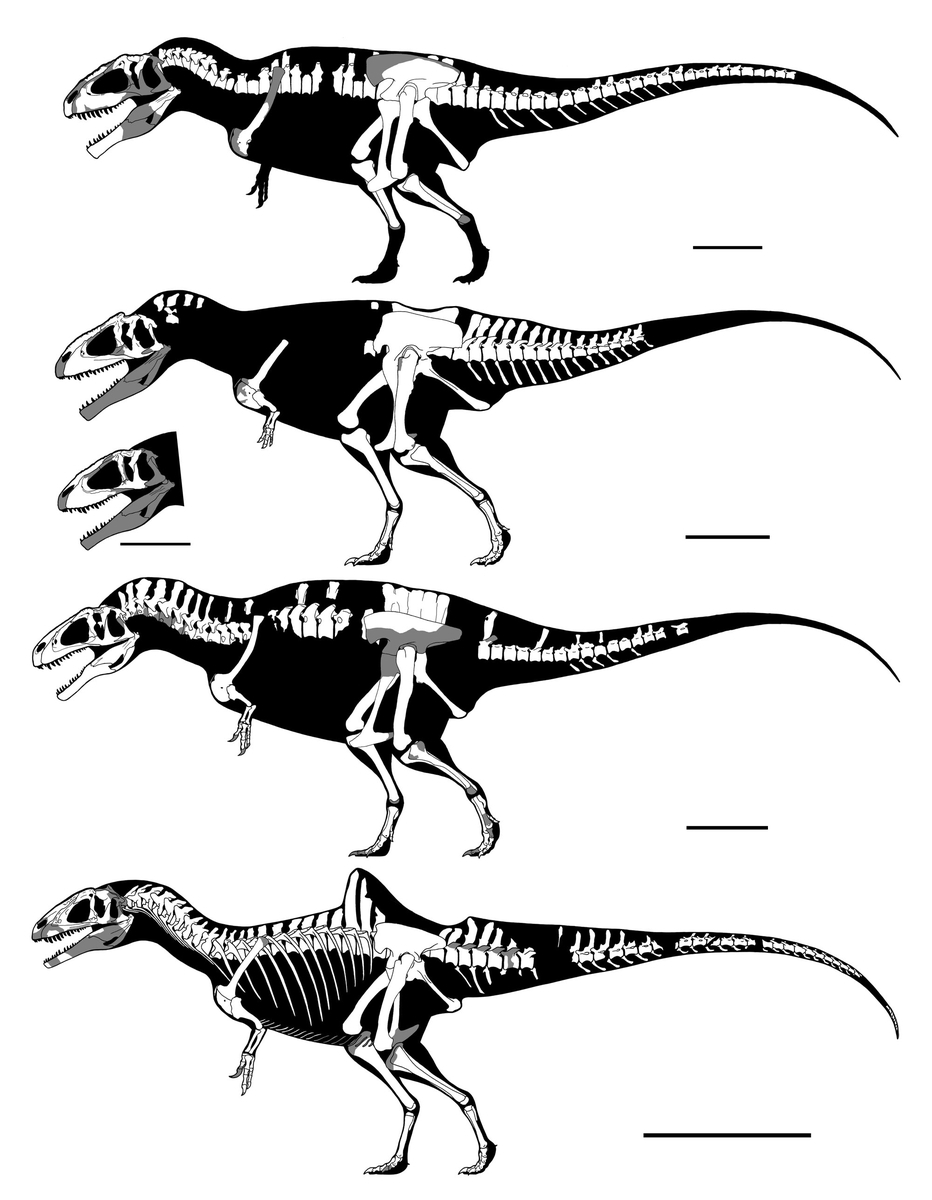
↑ティラノサウルス・レックス“スーSUE”FMNH PR2081(上)と“合衆国のT. レックス Nation’s T. rex(ワンケル・レックス)”USNM PAL 555000(下)。
スケールバーは1m。
“スー”はがっしり型の、“ワンケル・レックス”はきゃしゃ型の典型とされることがしばしばある。ポールは近年これらをホロタイプとしてティラノサウルス属に新種を設けたが、これは全く顧みられていない。
1980年代から90年代にかけて、ティラノサウルス・レックスの良好な骨格の発見ラッシュがあった。“ワンケル・レックス”ことMOR 555、“スー”ことFMNH PR2081――ラーソンは頑なに「旧標本番号」BHI-2033を併記し続けた――、そして“スタン”ことBHI-3033はいずれも数奇な運命を辿り、その他相当な数の部分骨格がプライベートコレクションへと消えていくこととなった。
1988年、モンタナはフォート・ペック貯水池(国立公園の一部、つまりは連邦政府の土地であり、陸軍工兵隊の管理下にあった)にて一家でボート遊びをしていたキャシー・ワンケルはダム湖に浮かぶ島へと上陸し、そこでいくばくかの化石を発見した。彼女はこれを最寄りの博物館――モンタナ州立ロッキーズ博物館(MOR)へと持ち込み、同定にあたったホーナーを仰天させた。これはティラノサウルスの前肢――記載されていたのは上腕骨だけだった――に違いなかったのである。所有権はあくまでも陸軍工兵隊にあったが、ともあれ発掘と(当分の間の)管理はMORが行うことになり、“ワンケル・レックス”――AMNH 5027をはるかに上回る完全度の骨格が姿を現した。頭骨は不完全であり、尾も前方1/3しか残っていなかったが、デスポーズでつながった椎骨の傍らには肩帯と完全な後肢があったのである。ワンケルが持ち込んだ前肢はほぼ完全であり、この骨格にはMOR 555のナンバーが与えられたのであった。
MOR 555のクリーニングが進められる一方で、サウスダコタでもそれを上回る発見があった。1990年、サウスダコタ各地の私有地をまわって販売用の化石を集めていたブラックヒルズ地質学研究所(BHI)が出くわしたのはほぼ完全かつすさまじく巨大で重々しいティラノサウルス・レックス――“スー”だったのである。これはやがてBHIを危機的状況に陥らせることになり、結局手元にも残らなかった――ラーソンはBHIの私設博物館でこれを展示するつもりだった――が、ほどなくして発見された“スタン”はBHIを救うことになった。ティラノサウルス・レックスの中でも最高の保存状態(かつ関節が完全に外れていたために内側面の観察も可能だった)とBHIが豪語する頭蓋を備えた“スタン”のキャストは世界中の博物館にあふれ、BHIを世界でも有数の化石業者へと押し上げたのである。“スタン”の実物復元骨格のこけら落としは日本で行われ、やがて“スー”の代わりにブラックヒルズの博物館のホールへ鎮座することになった。
紆余曲折の末にピーター・ラーソンは(“スー”のもろもろとは別件で)実刑に服し、そして1997年に競売にかけられた“スー”は(プライベートコレクションへ渡ることを危惧した末に、ディズニーとマクドナルド他の資金提供を受けた)FMNHによって落札された。手始めにカーペンターらによってMOR 555と“スー”の前肢が記載され、次いで2002年、ブロシュによって“スー”のモノグラフが出版されたのであった。
(“スー”と“スタン”の発見は、ティラノサウルスひいては恐竜の性的二形に関する研究に火を点けた。ティラノサウルスにおける“がっしり型”と“きゃしゃ型”はよく知られた話題であるが、これがピーター・ラーソンの言うように「二形」をなすのかは微妙なところである。加齢や個体変異の影響は排除できるものではなく、肯定的な評価は特に受けていない現状でもある。バッカーやラーソンはティラノサウルス・“エックス”――AMNH 5027などに代表されるもう一つの種が存在し、それらにも同様の性的二形がみられるとさえ述べたが、これも特に顧みられていない。)
幾度か論文にて記載されたのち、“スタン”を襲ったのはBHIを取り巻く法廷闘争であった。最終的に“スタン”は多くを語らぬまま裁判所の命令によって競売にかけられ、“スー”をはるかに上回る高額――化石の取引額としては断トツで最高額であり、もはや公的機関の手の届く額ではなかった――でどことも知れぬプライベートコレクションへと消えた。いつか夢見た詳細な骨学的記載はついぞなされることなく、BHIの希望の星として常に輝いていたはずの“スタン”は、キャストの販売権だけを残して展示ホールから去っていったのである。
ティラノサウルス(だけには当然とどまらないが)を取り巻くこうした状況は、カーをして「絶滅危惧種」と言わしめるものであった。“スー”並みの紆余曲折を経た“モンタナ闘争化石”――完全な前肢を保存した“ナノティラヌス”と、トリケラトプス・ホリドゥスと思しきほぼ完全らしい角竜――は(不発に終わった競売を経て)暗闘の末にノースカロライナ科学博物館の所蔵となったが、手放しで喜べる話なわけはない。暗がりの中の出来事が非公表なのはある種当然のことでもあるが、「重要な」標本の取引価格の高騰に歯止めがかからなくなっているのはもはや明らかなのである。「市場価値」と学術的なそれの間に絶望的な隔たりが生まれているのも違いなかった。前者を生み出すのは連綿と続く漠然としたティラノサウルスのイメージ――オズボーンの時代から喧伝されてきたそれが、研究を取り巻く状況を危機的なまでに追いつめていたのである。
(“スタン”は結局アブダビ自然史博物館に購入されていたことが判明したが、これが歓迎すべき事態かどうかは現状かなり微妙である。展示標本に対する研究のハードルは結局のところ非常に高い場合が多い。)
「絶滅危惧種」とはいえ、ティラノサウルスの復元骨格はわりあいに世界中どこでも見られるものである。AMNH 5027が担っていたティラノサウルスの「顔」としての役割はスタンへと引き継がれ、そしてそれに続く数々の新発見によって博物館で見られるティラノサウルスの「顔」はずいぶんにぎやかになった。アルバータと比してもまともな発見のなかったサスカチュワンはフレンチマンFrenchman層でもいきなり巨大なティラノサウルスの部分骨格――かなり完全な頭骨を保存していた――“スコッティ”ことRSM 2523.8が発見されたのである。一方で、コロラド以南のティラノサウルス・レックス(あるいはそれとごく近縁のもの)らしい化石は相変わらずお寒い状況でもあった。
AMNH 5027はリノベーションによって徹底的に修復されたのちに組み直され、今日もAMNHの顔役として来館者を出迎えている。CM 9380もリノベーションに際して解体され、頭骨要素は石膏の中から救出された(直後に来日した)のちに様々なティラノサウルスのキャストと組み合わされ、MOR 980――“ペックス・レックス”として知られる、MOR にとって2体目のかなり完全なティラノサウルス――のキャストとエドモントサウルスを奪い合う格好でマウントされた。オズボーンの果たせなかった夢が、AMNHのライバルであったカーネギー博物館によって(いささか形は変わったが)実現されたのである。“ディナモサウルス”のマウントは解体されて久しいが、それでもその立派な歯骨は展示され続けている。
長らくMORで産状風にマウントされ続けていたMOR 555のオリジナルは、2013年からスミソニアン――大規模なリノベーションを控えていた――へ(陸軍工兵隊によって)50年の期限で貸し出されることになった。名実ともに“合衆国のT. レックス”となったこの標本――新たにUSNM PAL555000(外部から受け入れたレプリカ等々にPALナンバーが付与される)の標本番号が与えられた――は、トリケラトプスの“ハッチャー”を供されてUSNMの新たな化石ホールに君臨することとなったのである。
1906年の暮れに展示デビューを果たしたティラノサウルスは、以来展示の王者として世界中の博物館に居座り続けてきた。周囲を取り巻く研究事情はダイナミックに動き続け、そして決して見通しの明るいものでもないのだが、ティラノサウルスたちは今日も玉座の上から、ちっぽけな人間たちのするさまを見下ろすだけである。


{kind=link}
{kind=link}
{kind=link}
/https://tf-cmsv2-smithsonianmag-media.s3.amazonaws.com/filer/20110520083220tyrannosaurus-mount-202x300.jpg){kind=link}
{kind=link}