「化石の日」である。こと日本においては今年が初めての試みではあるのだが、一応筆者も学界を追放されたわけではない(ちゃんと学会費は払っている)ので、便乗してもたぶん怒られることはないだろう。
言うまでもなく自☆演☆乙なわけだが、そういうわけで(論文がwebに出てからしばらく経つのだが――冊子体になったのはつい先日の話である)お付き合い願いたい。
上部白亜系はややもすれば意外なほど日本各地に点在しているのだが、そうはいってもたいがい露出は貧弱で、一般(相変わらず謎のフレーズである)の
知名度は皆無に等しいものも少なくない。そういう意味で
那珂湊層群は数少ない関東地方の上部白亜系として、地元――
茨城県ひたちなか市では多少の――「
白亜紀層」――として
知名度を得ていた
(かつてここには「白亜紀荘(国民宿舎)」があり、今日そこは「ホテルニュー白亜紀(市営)」(ネーミングに若干のいかがわしさを感じるが、何ということはない普通の観光ホテルである)となっている)。 那珂川の河口を挟んで南北10kmあまり、南は
大洗港から北は磯崎漁港(ここ数年で妙に
知名度の上がった
ひたち海浜公園(もともと陸軍の飛行場→わりかし最近まで
アメリカ空軍の悪名高き射爆場であった)は目と鼻の先である)までには、関東では珍しく
岩礁地帯が続いている。この
岩礁地帯(ともう少し陸側)は妙にややこしい地質からなり(付加体のそれとは比べ物にならないが)、それがそのまま
研究史となっている。そしてそのもっとも北側――4kmあまりの
岩礁をなすのが
那珂湊層群――カンパニアン後期~マーストリヒチアン前期(ざっと7500万~7000万年前あたりだろう)の海成層であった。
日本における地質学や古生物学の歴史が明治時代までさかのぼれるのは言うまでもないことで、大洗~磯崎まで続く
岩礁地帯(いちいち書くのも面倒なので以下
那珂湊-
大洗海岸とよぶ)の地質学研究のはじまりも明治の中ごろ――
1888年(ボーンウォーズ真っただ中の頃である)までさかのぼることができる。これは水戸周辺の地質図(20万分の1)だったのだが、この中で
那珂湊-
大洗海岸の
岩礁地帯はすべて第三系とされた。その後の研究でも(双葉に上部白亜系――双葉層群が認識される一方で)この一帯の地層は第三系とされ続けたのである。
(実のところ1926年に作成された20万分の1の地質図では
那珂湊-
大洗海岸のうち
那珂湊海岸の
岩礁地帯が下部白亜系とされたりもしたのだが、いかんせんこの地質図は説明書を欠いた作りで、かつ印刷された数もわずかだったため特に顧みられることはなかったらしい。今となっては根拠は完全に謎なのだが、どうも地層の固結度あたりに基づいていたフシがある。)
そんなこんなで戦後もこの一帯――まとめて湊層とよばれていた――は鮮新統とされていたのだが、実のところ露頭をよく観察してやるとそう単純な話ではなさそうであった。鮮新統とされていたものの「拾える」貝化石はどれも中新世のもののようであったし、走向・傾斜
からしてこの「湊層」のうちの上部(
那珂湊海岸に露出するもの)の下部(つまり湊層上部の下部)の方が湊層上部の上部よりも上位に来そうだったのである。
こうして色々と揺さぶりがかかっていた中、1954年5月16日に事態は急変した。一人の学生が「湊層上部の上部」から2つの
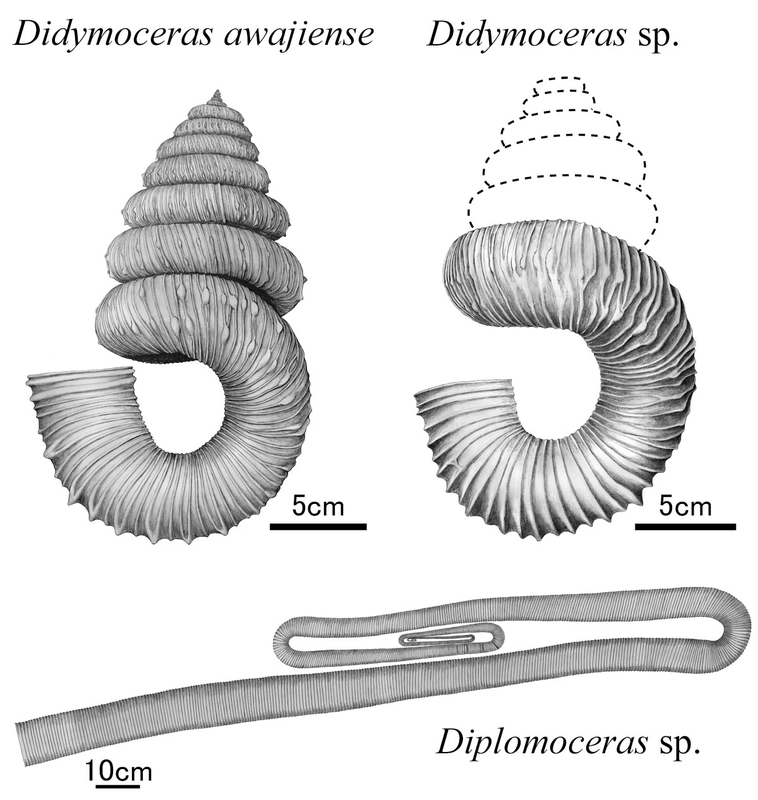
アンモナイト――ディディモセラスに似たフックの雌型と、バキュリテスを見出したのである。
このわずか2つ――前者(雌型とはいえ妙に保存がよい)はともかく後者の保存状態は悲惨だった――の化石が
那珂湊-
大洗海岸の地質を文字通りひっくり返すことになった。「湊層の下部」が上部白亜系大洗層に、「湊層上部の上部」が上部白亜系
那珂湊層(のち層群に格上げ)に、そして「湊層上部の下部」が中新統殿山層となったのである。
(この地から初めて
アンモナイトの化石を見出した学生は、そのまま大量の化石を採集(彼の調査ののち、これほど大量の化石がまとまって採れたことはとうとうなかった)するとともに一帯の地質を卒論としてまとめ上げた。その後研究者となることはなかったが、教員として
茨城県の地学教育普及に力を尽くしたという。)
大洗植物群の研究も含め、このあたりの地質と化石に関する研究は1960年代でひと段落付くことになった。
岩礁地帯に露出していたノジュールはあらかた採り尽くされ、それなりの数の軟体
動物化石――ほとんどが異常巻
アンモナイト――が採集されたのである(イノセラムスさえロクに出ず、そしてどういうわけか通常巻
アンモナイトは産出しなかった)。
異常巻
アンモナイトもいくらかはバキュリテスだったが、ほかはほとんどディディモセラス属――
D.アワジエンゼであった。他に2標本が新種――
D.ナカミナトエンゼとされ、他のいくつかの新種(たとえばブンブクウニ類――ニポナスター・ナカミナトエンシス)とともに「ご当地化石」の座に収まった。
やがて大洗層が白亜系から外され(どうも古第三系の気があるが、現状謎のままである)、
那珂湊層は
那珂湊層群に格上げされた(同時に部層が層へと格上げされ、下から順に築港層、平磯層、磯合層となった。築港層の露頭は今日完全に失われているのだが(ちょうど県
立海洋高校のあたりである)、そもそも磯合層の“切れ端”に過ぎない可能性さえある)。80年代に入り、北海道――
蝦夷層群に留まらず四国や淡路島、和泉山地――和泉層群における同時代層の
アンモナイトが続々と(再)記載された――が、
那珂湊層群は特段顧みられることはなかった。多様というわけでもなかったし、基本的に出てくる異常巻
アンモナイトは淡路島のものと変わり映えしなかったのである(保存もよくなかったし、そして何より地層としての規模がごく小さかった)。層序学的および堆積学的研究がぽつぽつと行われる一方で、
蝦夷層群や和泉層群を横目に古生物学的研究の進展はほとんどなかった。
状況が変わり始めたのは90年代に入ってからで、堆積相解析の概念を導入した再検討が(卒論ではあったが)おこなわれた。堆積学的な研究が中心であったとはいえ、通常巻
アンモナイトの破片とともに久方ぶりのみごとなディディモセラスの化石――GIUM 5001――が採集された(そしてその「雌型」は現地に残された;どうもこいつは新種くさいのだが、螺塔がきれいに失われており、また付近の層準からひとかけらたりとも参照できそうな化石が産出していないこともあってsp.止まりが無難なところである)。
それからの20年で少しずつ化石は増え(
茨城県自然博物館が開館したのも大きかった)、中には
那珂湊層群ではまだ報告されていなかったタイプのもの――クリップ状に巻くもの――ものも少なからず含まれていた。それまでアオザメ属の歯がただ一本発見されているだけだった
脊椎動物化石についても
翼竜の肩甲骨スッポン類の上腕骨やモササウルス類の尾椎が発見されるなど、着実に標本は増えつつあり、また(いかんせん狭いわりに開けて見通しの利く場所であり、産出地点はピンポイントで柱状図に落とし込むことができた)生層序的なデータの素も集まりつつあった。また、平磯層産の微化石の研究もおこなわれるようになった。
この頃になると、日本各地の上部白亜系で層序やらなんやらの再検討が続々と出版されるようになっていた。お膳立てはすべて整っていたのである。
役者がそろったのは2014年になってからで、GIUM 5001の雌型の「再」発見に始まり、
那珂湊層群産の異常巻
アンモナイトと各地の同時代層――和泉層群や
蝦夷層群産の標本との比較がおこなわれるようになった。同時に、溜まりに溜まった産地データは柱状図に叩き込まれ、1900mあまりに渡って途切れずに見える岩相の変化(すなわち堆積環境の変化である)と産出種の変遷が明るみに出た(意外なことに、先行研究で試みられたことはなかったのである)。
先行研究でそれとなく指摘されていたことではあるのだが、かくして、海底
扇状地の下部(平磯層下部)から中部~上部(平磯層上部~磯合層)へと堆積環境は移り変わり、そしてそれと合わせるように大型化石もディディモセラス主体からバキュリテス主体(そもそも平磯層下部とそれより上とでは化石の産出頻度が比べ物にならないのだが)へと移り変わっていくことが明白となった。平磯層の下部と磯合層とでは明らかに別の時代――カンパニアン後期とマーストリヒチアン前期――を示す化石が産出しており、そしてそれらは淡路島と北海道――堆積環境の違いから、(時代が被っているのは明らかであったにも関わらず)直接対比の困難だった一大産地を結び合わせるものでもあった。平磯層下部ではディディモセラス・アワジエンゼ――
那珂湊層群のほか四国と淡路島の和泉層群で多産する一方、北海道では未だ出る出る詐欺状態の種――が多産する一方、磯合層上部では「トゲの長い大型ノストセラス」――
那珂湊層群で最初に発見された
アンモナイトの片割れ――や“イノセラムス”・クシロエンシスといった、目下北海道(“
I.”クシロエンシスはサハリンやらでも出るのだが)でのみ知られているものと同様のものが産出していたのである。
那珂湊層群からこの60年あまりで出てきた異常巻
アンモナイトはいずれも殻のどこかしらを派手に欠いていたが、それでもいくつか淡路島や北海道のものに匹敵する(殻表面の保存やらに関していえばむしろ上回っているものもある)保存状態のものもあった。ディディモセラス・アワジエンゼの中には殻口が完全に保存されているものもあったし、ディプロモセラス(もろもろの理由でsp.だが、北西太平洋地域のものをひっくるめて究極的には新しい種小名が必要かもしれない)の中には――ものがものだけにポリプチコセラス属にされかけていた――成長中期の4本のシャフトが揃ったままのものさえあったのである。
(ディプロモセラスは世界中で産出し、日本でも中部カンパニアン以上でわりあいよく出るが、カンパニアン後期の後半――ディディモセラス・アワジエンゼと同じ層準から出てきたケースはそれまで未報告であった。また、シャフトが化石化の過程でばらけることがよく知られており(成長後期の殻が2本つながっていればいい方である)、成長中期の殻が4本まとまって出てくるケースは極めて稀である。こうした事情から、その辺でみかけるディプロモセラスの復元図は成長初期~中期の殻を文字通り端折られていたりする。)
この60年で海岸沿いの風景は激変し、ディディモセラス・ナカミナトエンゼもD.アワジエンゼのシノニムとなったが、それでも岩礁地帯は、そして海は――7500万年前から――変わらずそこにある。どうにか連綿と“バトン”として受け継がれてきた標本たちは今日も新たな研究者の訪れを待っており、そして潮が引くたび那珂湊層群の化石は――筆者の見つけ損ねたものたちは細粒タービダイトの波食台の上で、太陽に身を焦がしている。
(シロワニ属の歯や“ヒタチナカリュウ”として知られるニクトサウルス類の肩甲骨巨大スッポン類の上腕骨、モササウルス類の尾椎、そして推定甲長80cmに達する巨大なスッポン類の背甲の産出した層準は、どうもマーストリヒチアン初頭――淡路島のランベオサウルス類やカムイサウルスと同時代に相当するようだ。もし――もしも恐竜が出てくるとすれば、たぶんこの層準を置いて他にない。)