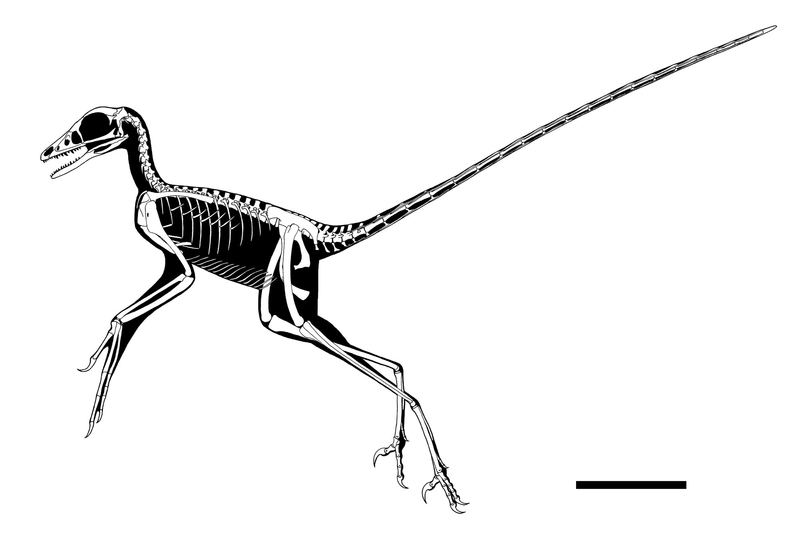
Composite skeletal reconstruction of Microraptor zhaoianus.
Largely based on IVPP V13352 (holotype of M. "gui") .
Some details reconstructed after BMNHC PH881, CAGS 20-7-004, CAGS 20-8-001, and LVH 0026 (holotype of M. "hanqingi").
Scale bar is 10cm.
このところ微妙に忙しい日が続いたりもして(意外とそうでもなかったかもしれない)ブログの更新がほったらかされていたわけである。悪だくみの成果はとりあえず実を結びそうで、あるいはそのうちなにがしかの発表ができるかもしれない。
ミクロラプトルといえば命名当初(むしろそれ以前)から高い知名度を誇っていたわけだが(断言)、羽毛に注目こそされど、骨格のつくりやら何やらに関しては意外なほど知られていない。まとまった数の全身骨格が出ている一方で、骨そのものの保存のよい化石は少なく、そのあたりが障害となっていたわけである(記載そのものはわりあい積極的に行われていたりもしたのだが)。それでもここ数年で状態のよい標本の記載が進んだこともあり、真面目に骨格図を描けるような状態に至ったといえる。そういうわけで、ミクロラプトルについて今回も適当に書いておきたい。
遼寧省の下部白亜系(00年代初頭まで上部ジュラ系とみなす意見もあったわけだが)といえばそれなりに古くから極めて保存のよい化石で知られていたのだが(時代が時代だったので、軒並み日本人研究者によって記載された)、羽毛を保存した化石が本格的に産出するようになったのは90年代あたりからだったりする。孔子鳥Confuciusornisに代表されるおびただしい数の鳥類化石が産出するようになると、瞬く間に中国国外向けの輸出市場が―――ブラックマーケットが形成されたのだった。
さて、ミクロラプトルの模式標本となる化石は1997年に地元住民によって発見され、そして数奇な運命を辿ることとなった。メインパートはIVPPへ売られた一方で、カウンターパートは鳥類化石と(その辺の適当な岩くずも混ぜつつ)合体させられ、そのまま(密輸された末に)ツーソンでツェルカスの手に渡ることとなったのである。
ツェルカスの手に渡ったこのアーティファクトの顛末についてはここで書くこともないだろう。かくしてすったもんだの末に(アルカエオラプトルのシノニムになりかけた末に)カウンターパートの下半身部分(とIVPPの購入した上半身の要素を含むメインパートIVPP V 12330)は徐星らによって新属新種の基盤的小型ドロマエオサウルス類ミクロラプトル・ジャオイアヌスMicroraptor zhaoianusと命名されたのだった。ナショナルジオグラフィックの特報から1年後、2000年のことである。
(“アルカエオラプトル”の上半身はその後2002年になって、よりによってツェルカスと徐星の共著でアルカエオヴォランスArchaeovolansと命名された。が、その年のうちにヤノルニスYanornis―――命名は2001年―――のシノニムとなってしまった。)
ミクロラプトル・ジャオイアヌスの模式標本IVPP V 12330は下半身こそ羽毛と共によく保存されていたものの、上半身については部分的な前肢と胴体の一部、原形を留めないレベルで潰れた頭骨しか残っていなかった。とはいえ、2002年になってほぼ完全な骨格が記載されることとなった。
2003年にミクロラプトル属の新種ミクロラプトル・グイMicroraptor guiのホロタイプとして記載された全身骨格IVPP V13352は驚くべきことに前肢だけでなく後肢にまで長い羽毛―――左右非対称の風切羽―――を保存しており、鳥類の飛翔の起源に関する議論に一石を投じることとなった。四肢に翼をもつミクロラプトル・グイは、1915年にビーベが理論上の存在として示した“テトラプテリクス段階Tetrapteryx stage”(余談だが学名としてのTetrapteryxも存在しており、こちらはハゴロモヅルのシノニムとなっている)そのものだったのである。
M.グイが話題をさらう一方で、2002年にはM.ジャオイアヌスの見事な骨格2体も記載されていた。これらの標本は頭骨こそ部分的だった(羽毛も残っていなかった)もののそれ以外の要素はよく保存されており、ミクロラプトル属の骨学的情報がかなり明らかとなった。一方で、依然として頭骨の詳細ははっきりしなかった(IVPP V13352も後頭部はよく残っていた(吻は原記載の時点で「後付け」であることが指摘されている)ものの、骨そのものの保存は全体としてよくなかった)。
その後もミクロラプトルに関する研究が盛んに行われたが、飛行能力に関するものがほとんどであった。食性に関する研究(樹上性の哺乳類やら鳥やら魚やら、とりあえず食べられそうな動物は何でも食べていたようだ)や羽毛に残されたメラノソームの研究なども行われる一方で、骨学的な記載はなかなか進まなかったのである。
とはいえ、ドロマエオサウルス科のレビューの中でM.グイとM.ジャオイアヌスの間に分類学的に有意な形態差がないらしいことが指摘されるなど、徐々にではあるが骨学的な情報に再びスポットが当たるようになった。2014年に凄まじく状態のよい標本が記載され、ここに至ってミクロラプトルの全身の骨格が明らかになったのである。
どさくさに紛れて命名されたクリプトヴォランスCryptovolans(ツェルカスは妙にvolansという言葉にこだわりがあったらしい)やM.ハンチンギhanqingiまでシノニムとして取り込んだ結果、ミクロラプトル・ジャオイアヌスの全長は結局1mまで「拡大」することになった。当初全長3~40cmの「最小の恐竜」とうたわれたミクロラプトルだったが、ふたを開けてみれば(依然として小型恐竜であることに違いはないが)ほどほどの大きさの生き物だったのである。
しばしば「小顔」として描かれがちだが、ミクロラプトルの頭部は体のサイズの割に大きく、かつそこそこ大きな歯が生えている。先述した小さな哺乳類(全長10cm弱)や鳥を捕食するのには十分すぎる武器といっていいのだろう。首が比較的短く、かつ胴体が長いのも割と見落とされがちな特徴である。
前肢は長い―――が、後肢が凄まじく長いので、全体のバランスとしては実のところそれほど長いものでもない。後肢の第I指が対向していたのかどうか確認する術は現状ない(ぺしゃんこに押しつぶされた化石でそういう話を議論するのはそもそも酷である)が、長さは非常に短く(このあたり、相対的に割と長い第I指をもつデイノニクスとは対照的である)、対向していたとしても木の枝を掴む役には立たなさそうである。一方で前肢の末節骨は後肢の「シックルクロー」と同サイズであり、四肢を使って木の幹に抱き着いていたというのがありそうな話である。尾は長いが細く、上半身のカウンターウェイトにはならなさそうだ。
腰帯はドロマエオサウルス科の典型―――からはやや外れて、恥骨には独立したブーツをもたない。恥骨のシャフトは途中でいきなり後方にへし折れるという(ミクロラプトル類に共通する)独特のつくりである。股関節は結局のところ(例えばデイノニクスと比べて)特殊化しており、多少なりとも外側に広げやすくはなっていたようだ。
ミクロラプトルの標本は地元民によって採集されたのち博物館に売却されたものが少なからずあり、従ってその産出層や時代については少々厄介なところがある。とはいえ、そのあたりのデータがはっきりしている標本はいずれも熱河Jehol層群の九佛堂Jiufotang層(白亜紀前期アプチアン;ざっと1億2100万年前ごろ)から産出したものである。
ドロマエオサウルス科の基盤的な位置に置かれているミクロラプトル類microraptoriaはシノルニトサウルス(結局のところミクロラプトルより気持ち大きい程度の計算になる)などいくつかの属を含むが、ほとんどは白亜紀前期後半かつ遼寧省産のものである。一方で、カナダはアルバータ州のダイナソー・パークDinosaur Park層(詳しい層準ははっきりしないが、とりあえずおおざっぱに7700万~7500万年前ごろか)からはヘスペロニクス―――実質的には腰帯が知られているだけだがミクロラプトルに酷似―――が産出していたりもする。いかんせん腰帯だけではあるが、ミクロラプトル類の恥骨は極めて特徴的な形態であり、ヘスペロニクスは一般にミクロラプトル類として扱われている。